
A BURGONYA Y VÍRUS KÖPENYFEHÉRJE GÉN ÁLTAL INDUKÁLT...
126
Szent István Egyetem Gödöllő A BURGONYA Y VÍRUS KÖPENYFEHÉRJE GÉN ÁLTAL INDUKÁLT REZISZTENCIA KIALAKÍTÁSA BURGONYA- ÉS DOHÁNYFAJTÁKBAN, ÉS EZEK SZÁNTÓFÖLDI ÉRTÉKELÉSE Doktori értekezés JÓZSA RITA Gödöllő 2002
Transcript of A BURGONYA Y VÍRUS KÖPENYFEHÉRJE GÉN ÁLTAL INDUKÁLT...

Szent István Egyetem
Gödöllő
A BURGONYA Y VÍRUS KÖPENYFEHÉRJE GÉN ÁLTAL
INDUKÁLT REZISZTENCIA KIALAKÍTÁSA BURGONYA- ÉS
DOHÁNYFAJTÁKBAN, ÉS EZEK SZÁNTÓFÖLDI
ÉRTÉKELÉSE
Doktori értekezés
JÓZSA RITA
Gödöllő
2002

2
A doktori iskola neve: Biológiai Doktori Iskola
Tudományága: Biológiatudomány
Vezetője: Dr. Tuba Zoltán
tanszékvezető egyetemi tanár
Szent István Egyetem, Mezőgazdaság- és
Környezettudományi Kar, Növénytani és
Növényélettani Tanszék
Témavezető: Dr. Balázs Ervin
akadémikus, intézetigazgató
Mezőgazdasági Biotechnológiai Kutatóintézet,
Környezet Biotechnológiai Intézet
.................................... ....................................Dr. Tuba Zoltán Dr. Balázs Ervin
a doktori iskola vezetője témavezető

3
TARTALOMJEGYZÉK:
RÖVIDÍTÉSEK JEGYZÉKE: ..................................................................................................6
1. BEVEZETÉS ..........................................................................................................................8
2. IRODALMI ÁTTEKINTÉS ................................................................................................11
2.1. A BURGONYA Y VÍRUS (PVY) JELENTŐSÉGE......................................................11
2.2. A PVY RENDSZERTANI BESOROLÁSA ÉS JELLEMZÉSE....................................13
2.3. PATOGÉNTŐL SZÁRMAZTATOTT REZISZTENCIA..............................................18
2.3.1. Klasszikus keresztvédettség .....................................................................................18
2.3.2. Molekuláris módszerekkel kiváltott ellenállóképesség ............................................202.3.2.1. Köpenyfehérje (CP) jellemzése ..................................................................................... 21
2.3.2.2. A köpenyfehérje gén által közvetített rezisztencia (coat protein mediated resistance -
CPMR)........................................................................................................................................ 23
2.3.2.3. Köpenyfehérje által közvetített keresztvédettség ........................................................... 24
2.3.2.3.1. Potyvirus nemzetség .............................................................................................. 25
2.3.2.4. A köpenyfehérje gén által indukált rezisztencia lehetséges hatásmechanizmusai.......... 29
2.3.2.4.1. A köpenyfehérje által közvetített rezisztencia hatásmechanizmusai...................... 30
2.3.2.4.2. Az RNS-ek által közvetített rezisztencia hatásmechanizmusai, a géncsendesítés
jelensége................................................................................................................................ 32
2.3.2.5. Kockázati tényezők a köpenyfehérje által mediált keresztvédettség technológiájának
felhasználása esetén .................................................................................................................... 38
2.3.2.5.1. Hetero- és transzkapszidáció ................................................................................. 39
2.3.2.5.2. Homológ és heterológ rekombináció ..................................................................... 41
2.3.2.5.3. Szinergizmus ......................................................................................................... 42
3. ANYAG ÉS MÓDSZER......................................................................................................43
3.1. ANYAGOK ....................................................................................................................43
3.1.1. Növényi anyag.........................................................................................................43
3.1.2. Vírusizolátum ..........................................................................................................44
3.1.3. Génkonstrukció .......................................................................................................44
3.1.4. Baktérium törzs .......................................................................................................45
3.1.5. Táptalaljok, tápoldatok ...........................................................................................46

4
3.1.6. Oldatok, pufferek.....................................................................................................47
3.2. MÓDSZEREK ................................................................................................................48
3.2.1. Módszerek transzgénikus növények előállításához..................................................483.2.1.1. Növény transzformáció.................................................................................................. 48
3.2.1.2. Hajtásindukálás.............................................................................................................. 49
3.2.1.3. Gyökereztetés ................................................................................................................ 49
3.2.1.4. Kiültetés üvegházba....................................................................................................... 49
3.2.2. Transzgénikus növények vizsgálata.........................................................................503.2.2.1. A köpenyfehérje gén integrációjának vizsgálata............................................................ 50
3.2.2.2. Transzgén expresszió vizsgálata .................................................................................... 51
3.2.2.3. Rezisztenciavizsgálatok ................................................................................................. 52
3.2.2.3.1. A mesterséges fertőzés és körülményei ................................................................. 52
3.2.2.3.2. Szabadföldi kísérletek............................................................................................ 53
3.2.2.3.2.1. Kisparcellás, infektor növényes, szabadföldi kísérlet .................................... 55
3.2.2.3.2.2. ELISA vizsgálat ........................................................................................... 57
3.2.2.3.2.3. Solanum demissum A6 teszt ......................................................................... 58
3.2.2.3.2.4. Kertészeti oltás kísérletek.............................................................................. 58
3.2.3. Vírusmentesítés .......................................................................................................59
3.2.4. Vírustisztítás............................................................................................................59
4. EREDMÉNYEK ...................................................................................................................60
4.1. TRANSZGÉNIKUS NÖVÉNYEK ELŐÁLLÍTÁSA ....................................................60
4.2. TRANSZGÉNIKUS NÖVÉNYEK VIZSGÁLATA ......................................................62
4.2.1. Az integrálódott CP gén kimutatása........................................................................62
4.2.2. Transzgén expresszió vizsgálata .............................................................................63
4.2.3. Rezisztenciavizsgálat üvegházban, dot-blot analízis ...............................................65
4.2.4. A molekuláris vizsgálatok eredményeinek összegzése.............................................69
4.2.5. Szántóföldi vizsgálatok............................................................................................714.2.5.1. Dohányvonalak szabadföldi vizsgálata .......................................................................... 71
4.2.5.2. A transzgénikus burgonyavonalak szabadföldi vizsgálata ............................................. 73
5. EREDMÉNYEK MEGVITATÁSA, KÖVETKEZTETÉSEK.........................................85
6. ÖSSZEFOGLALÁS, ABSTRACT......................................................................................90
7. IRODALOM .........................................................................................................................92

5
A DISSZERTÁCIÓHOZ KAPCSOLÓDÓ TUDOMÁNYOS KÖZLEMÉNYEK
JEGYZÉKE: ...........................................................................................................................108
KÖSZÖNETNYILVÁNÍTÁS................................................................................................110
1. MELLÉKLET ...............................................................................................................111
ALFAMOVIRUS NEMZETSÉG....................................................................................................111
CARLAVIRUS NEMZETSÉG.......................................................................................................112
CUCUMOVIRUS NEMZETSÉG...................................................................................................112
ENAMOVIRUS NEMZETSÉG .....................................................................................................113
FUROVIRUS NEMZETSÉG........................................................................................................113
GEMINIVIRUS NEMZETSÉG .....................................................................................................114
ILARVIRUS NEMZETSÉG..........................................................................................................114
LUTEOVIRUS NEMZETSÉG ......................................................................................................114
NEPOVIRUS NEMZETSÉG ........................................................................................................115
POTEXVIRUS NEMZETSÉG ......................................................................................................115
TENUIVIRUS NEMZETSÉG .......................................................................................................116
TOBAMOVIRUS NEMZETSÉG ...................................................................................................117
TOBRAVIRUS NEMZETSÉG ......................................................................................................118
TOMBUSVIRUS NEMZETSÉG....................................................................................................119
TOSPOVIRUS NEMZETSÉG ......................................................................................................119
2. MELLÉKLET ...............................................................................................................122
KÖPENYFEHÉRJE GÉNEK ÁLTAL KIVÁLTOTT REZISZTENCIA VÍRUSNEMZETSÉGENKÉNT ...............122

6
RÖVIDÍTÉSEK JEGYZÉKE:
BAP benzil-aminopurinBA benzil-adeninbp base pair, bázispárCCC klór-kolin-kloridcDNS copy/komplementer DNSCP coat protein, köpenyfehérjeCPMR coat protein mediated resistance, köpenyfehérje által közvetített rezisztenciadpi days past inoculation, a fertőzést követő x. napondsRNS double stranded, dupla szálú RNSELISA enzyme linked immunosorbent assayHDR homology dependent resistance, homológiafüggő rezisztenciaH 11 Hevesi 11 dohányfajtaIAA indol-ecetsavM Mindenes burgonyafajtaNAA naftil-ecetsavNCR non coding region, nem kódoló régióPCR polymerase chain reaction, polimeráz láncreakcióPDR pathogen derived resistance, patogéntől származtatott rezisztenciaPTGS post-transcriptional gene silencing, poszt-transzkripcionális géncsendesítésSK Somogyi Kifli burgonyafajtassRNS single stranded, egyszálú RNSSC Stamm C2 dohány nemesítési vonalVLP virus like particulum, vírus-szerű parikulumVD Virgin D dohányfajta
Vírusok:
AMV alfalfa mosaic virus, lucerna mozaik vírusArMV arabis mosaic virus, arabis mozaik vírusBMV brome mosaic virus, rozsnok mozaik vírusBNYVV beet necrotic yellow vein virus, répa nekrotikus sárgaerűség vírusBYMV barley yellow mosaic virus, árpa sárga mozaik vírusCaMV cauliflower mosaic virus, karfiol mozaik vírusCMV cucumber mosaic virus, uborka mozaik vírusCyMV cymbidium mosaic virus, cymbidium mozaik vírusCyRSV cymbidium ringspot virus, cymbidium gyűrüsfoltosság vírusINSV impatiens necrotic spot virus, nebántsvirág nekrotikus foltosság vírusGBNV groundnut bud necrosis virus, földimogyoró rügynekrózis vírusGCMV grapevine chrome mosaic virus, szőlő króm mozaik vírusGRSV groundnut ringspot virus, földimogyoró gyűrűsfoltosság vírusMDMV maize dwarf mosaic virus, kukorica törpe mozaik vírusPAMV potato aucuba mosaic virus, burgonya aucuba mozaik vírusPEMV pea enation mosaic virus, borsó enációs mozaik vírusPepMoV pepper mottle virus, paprika foltosság vírus

7
PLRV potato leaf roll virus, burgonya levélsodródás vírusPPV plum pox virus, szilva himlő vírusPRSV papaya ringspot virus, papaya gyűrüsfoltosság vírusPStV peanut stripe virus, földimogyoró csíkosság vírusPVM potato M carlavirus, burgonya M vírusPVS potato S virus, burgonya S-vírusPVX potato X virus, burgonya X vírusPVY potato Y virus, burgonya Y vírusRSV rice stripe virus, rizs csíkosság vírusSMV soybean mosaic virus, szója mozaik vírusSHMV sunn-hemp mosaic virus, kender mozaik vírusTCSV tomato chlorotic spot virus, paradicsom klorotikus foltosság vírusTEV tobacco etch virus, dohány karcolatos vírusTMV tobacco mosaic virus, dohány mozaik vírusToMV tomato mosaic virus, paradicsom mozaik vírusTRV tobacco rattle virus, dohány rattle vírusTSWV tomato spotted wilt virus, paradicsom foltos hervadás vírusTSV tobacco streak virus, dohány csíkosság vírusTuMV turnip mottle virus, tarlórépa tarkulás vírusTVMV tobacco vein mottling virus, dohány érfoltosság vírusTYLCV tomato yellow leaf curl virus, paradicsom sárga levélgöndörödés vírusWMV-2 watermelon mosaic virus-2, görögdinnye mozaik vírusZYMV zucchini yellow mosaic virus, cukkíni sárga mozaik vírus

8
1. BEVEZETÉS
Gazdasági növényeink termőképességének növelése vagy szinten tartása
szorosan összefügg a növények betegségekkel, kórokozókkal szembeni
ellenállóképesség fokozásával. A növénytermesztésben jelentős károkat okozó
növényi vírusok ellen a mezőgazdaság szakemberei mindig is alkalmaztak
különböző módszereket és alkalmaznak ma is. A védekezés korábban főként
preventív jellegű volt, emellett intenzív nemesítő munka folyt és folyik jelenleg
is a növények vírus-ellenállóságának kialakítása terén. A vad fajokban (pl.
Solanum stoloniferum, S. brevidens) meglévő rezisztenciagének többnyire más
tulajdonsággal kapcsoltak, így ezek felhasználása csak hosszadalmas és
munkaigényes folyamat után válik lehetővé, ráadásul ezek a gének egy erősebb
vírusfertőzés esetén már nem nyújtanak megfelelő védelmet.
A vírusok által okozott károk elsősorban járványok megjelenésekor érnek
el szembetűnő gazdasági veszteségeket. Világszerte komoly gondot okoz
főképpen a burgonya- és dohánytermesztésben a burgonya Y vírus (PVY)
kártétele. A vírus nemcsak a terméshozam csökkentésével idéz elő
veszteségeket, hanem ezáltal értékes és kiváló tulajdonságokkal rendelkező régi
magyar fajtákat kiszorít a köztermesztésből. Ezek a fajták a vírussal szembeni
fogékonyságuk következtében a növénytermesztés számára jelenleg
használhatatlanok, így csupán génbankokban találhatóak meg. Ennek
következtében jelentős mértékben csökken a mezőgazdaság biodiverzitása.
Az 1970-es éveket követően Magyarországon a burgonya leromlása
jelentős méreteket öltött, így az államilag elismert fajták, mint például a
Gülbaba, az Aranyalma, a Kisvárdai Rózsa vagy a Mindenes a
köztermesztésből kiszorultak. A Hollandiából importált fajták, mint pl. a
Desiree, vetőgumócserés rendszerével a hazai fajták nem tudtak versenyezni. A

9
leromlásra hajlamosító tényezők Magyarországon elsősorban a PLRV (potato
leafroll virus), az Y, X, A, S burgonyavírusok, valamint más kórokozók közül a
fitoftóra és az alternária, így az ezekkel szembeni jó szántóföldi rezisztenciájú
fajták előállítása vált fontossá. A fitoftórával illetve az alternáriával szembeni
védekezés vegyszeresen részben megoldott, viszont a vírusokkal szemben a
rezisztencianemesítés illetve a génsebészeti módszerek a leghatékonyabbak. A
levéltetűvektorok elterjedését ugyan vegyszeres védekezéssel vissza lehet
szorítani, azonban a leghatékonyabb megoldás mégis a vírusrezisztens
transzgénikus növények előállítása, melyek felhasználásával csökkenthető a
vírusvektorok ellen alkalmazott inszekticidek mennyisége is és megvalósítható
egy környezetkímélőbb növényvédelem.
A nyolcvanas években terjedt el hazánkban egy új PVY törzs, mely a
korábbi rezisztens burgonyafajtákat is képes volt megbetegíteni (Beczner és
mtsai, 1984). Ez a vírustörzs rezisztencia-áttörő képességgel rendelkezik (Le
Romancer-Kerlan, 1992., Van den Heuvel et al., 1994) és jellegzetes nekrotikus
tüneteket okoz a gumókon. A burgonya gumó nekrotikus gyűrűsfoltosság
(PVYNTN) nevet kapta. Jelenleg az egész világon elterjedt és károsít.
Mára már két évtized telt el azóta, hogy géntechnológiai módszerekkel
valamely élő szervezetből tetszőlegesen kiválasztott tulajdonságokat hordozó
gének izolálására van lehetőség. Ezen gének egy másik szervezetbe történő
működőképes beépítésével a kívánt tulajdonság megnyilvánítható és örökíthető
(Balázs és Gáborjányi, 1984). Az így előállított transzgénikus növények (pl.
toleráns vagy rezisztens fajták) előállítása költségkímélőbb és időtakarékosabb,
mint a hagyományos módszerekkel nemesített fajtáké (Tepfer és Balázs, 1997).
A kórokozók újonnan megjelenő törzsei, biotípusai vagy rasszai azonban még
így is folyamatosan új kihívást jelentenek a szakemberek számára.

10
A genetikailag módosított növényekkel történő kutatásokat, szántóföldi
vizsgálatokat, engedélyeztetésüket és a köztermesztésbe kerülésüket törvények
szabályozzák (Balázs, 1997).
Az elmúlt közel két évtizedben a molekuláris biológiai eszközök
felhasználásával a világ különböző laboratóriumaiban vírusellenálló növényeket
hoztak létre. A víruseredetű géneket transzgénként felhasználó módszerek közül
legelterjedtebben alkalmazott a köpenyfehérje gént tartalmazó konstrukciókkal
történő növényi transzformáció, melyet számos növény-vírus kombinációban
használtak már sikerrel. Ezzel a módszerrel váltottak ki vírus-ellenállóságot
más jól ismert burgonyafajtáknál is, mint pl. Russet Burbank, Bintje és Desiree.
Munkánk során célul tűztük ki, hogy a hazai flórából származó burgonya Y
vírus köpenyfehérje génjét a vírussal szemben fogékony, régi magyar
burgonya- és dohányfajtákba építsük be és az így létrehozott transzgénikus
növények által új, vírusellenálló vonalakat alakítunk ki. Célul tűztük ki azt is,
hogy az így előállított vonalak közül molekuláris- és előzetes
rezisztenciavizsgálatok által kiválasztott legkiválóbb vonalakat szántóföldi,
propagatív körülmények között is vizsgáljuk és bebizonyítsuk, hogy ezzel a
módszerrel előállított transzgénikus növények nagy hatékonysággal és
biztonsággal használhatóak a növénytermesztésben, visszaadva a rég elfeledett,
egykor kiváló fajtákat a mezőgazdaság számára.

11
2. IRODALMI ÁTTEKINTÉS
2.1. A BURGONYA Y VÍRUS (PVY) JELENTŐSÉGE
A zöldség és dísznövények egyik gazdaságilag legjelentősebb
víruskórokozója a PVY, mely főképpen a Solanaceae családba tartozó
növényeket károsítja. Jelenlegi ismereteink szerint 405 növényfajt fertőz 31
családon és 72 nemzetségen belül (Edwardson és Christie, 1997). Megfertőzi a
burgonya, dohány (1., 2. ábra), paradicsom, paprika kultúrákat, és néhány
gyomot is, mint például a Solanum nigrum L., Portulaca oleracea L. A PVY a
burgonya legfontosabb és legsúlyosabb károkat előidéző vírusbetegségének a
kórokozója, mely minden burgonyatermesztő országban előfordul. A vírus a
burgonya X (PVX) és a burgonya levélsodródás vírussal (PLRV) együtt okozza
a burgonya vírusos leromlását, amely így akár 10-80%-os terméskiesést is
eredményezhet (Hollings és Brunt, 1981, Horváth, 1995).
1. ábra PVY által okozott tünetek dohánylevélen.

12
2. ábra A PVY kártétele dohánytermesztésben.
(Debrecen-Pallag)
A PVY levéltetvek által terjed, nem perzisztens módon, azaz terjedése
gyors, de csak néhány óráig. Ellentétben a perzisztens módon terjedő
vírusokkal, mint pl. a PLRV, amely több napig a levéltetvek nyálmirigyében
maradva fertőz. Egyetlen fertőzőképes levéltetű több növényt is képes
megfertőzni. A vírus átvitelére mintegy 20 különböző nemzetségbe tartozó
levéltetű faj képes. A PVY a parenchimában lokalizálódó vírusok közé tartozik.
A PVYNTN legfontosabb természetes gazdanövénye a burgonya (Solanum
tuberosum ssp. tuberosum). Az elsődleges fertőzés általában kevésbé súlyos
következményekkel jár, mint a másodlagos, amely a burgonyagumókban
megmaradó vírusokból indul ki (3. ábra).

13
3. ábra
PVY fertőzési ciklus burgonyánál
(Farinelli, 1992).
A vírusterjedés függ a korai levéltetű-populációtól és a fertőzött növények
mennyiségétől. Az egészséges növények más növényfajokról, de gyakran a
fertőzött gumókból kihajtó növényekről kapják meg a vírust a levéltűvektorok
által. Fertőzött gumó pedig fennmaradhat a talajban átvészelve a telet, de az is
előfordulhat, hogy betakarítás után nem vehető észre a fertőzöttség és
felhasználják vetőburgonyaként a gumókat a következő évben.
2.2. A PVY RENDSZERTANI BESOROLÁSA ÉS JELLEMZÉSE
A Potyviridae család a nevét a PVY-ról kapta. A vírust először a XX.
század első felében Smith (1931) írta le Nagy-Britanniában. Majd Harrison és
Roberts (1971) sorolta a PVY-t és a hozzá hasonló vírusokat egy csoportba és
később a legjellemzőbb tagjáról nevezték el Potyvírus nemzetségnek. A
legnépesebb és gazdaságilag a legjelentősebb víruscsalád. Több, mint 200 tagja

14
van öt nemzetségen belül: Potyvírus-, Bymovírus-, Rymovírus-, Ipomovírus- és
Macluravírus nemzetség. (Shukla és mtsai,1994).
A potyvírusok hajlékony, fonal alakú növényi vírusok, átmérőjük 11-15 nm
hosszúságuk 650-900 nm, és megközelítőleg 5% nukleinsavat és 95% fehérjét
tartalmaznak (Dougherty és Carrington,1988). A virionok egyszálú, pozitív,
szensz RNS-t tartalmaznak, amely megközelítőleg 10.000 nukleotidból áll. A
potyvírusok genomszerkezetének pontosabb feltárása a vírusok nukleotid
szekvenciájának ismeretével és a vírus RNS komplementer DNS-éről (cDNS)
szintetizálódó fertőzőképes transzkriptumok létrehozásával vált lehetővé. A
vírust burkoló köpenyfehérje 2000 alegységből áll össze. Az RNS genom 5'
végéhez egy fehérje kötődik, a 3' végen pedig poliadenilált szekvencia található,
melynek hossza a vírus fertőzőképességét befolyásoló tényező (Riechmann és
mtsai, 1990). A potyvírus nemzetség tagjai és az egyes vírusok izolátumai
között nagy különbségek vannak a 3’ vég méretében és szekvenciasorrendjében
(Laín és mtsai, 1988, Dolja és Carrington, 1992). A nemkódoló régiók közül az
5’ vég sokkal konzervatívabb, mint a 3’ vég, így pl. az egyes víruscsaládokat a
3’ vég bázissorrendje alapján lehet megkülönböztetni (Atreya, 1992). A genomi
RNS-ről egy nagy poliprotein prekurzor (340-368 kDa) keletkezik (Allison és
mtsai, 1986), amelynek hasítása vírus által kódolt proteázokkal történik. A
poliprotein prekurzor hasítási termékei az N-terminális végtől a C-terminális
vég felé haladva a következők: P1- első fehérje, HC-Pro- segítő komponens-
proteáz, P3-harmadik fehérje, 6K1- feltételezett első 6 kDa-os fehérje, CI-
henger alakú zárványfehérje, 6K2- második 6kDa-os fehérje, VPg-NIa- kis
nukleáris zárványfehérje, NIb- nagy nukleáris zárványfehérje, valamint a CP-
köpenyfehérje. (Riechmann és mtsai,1992), (4. ábra).

15
4. ábra
Potyvírusok genomszerveződése és a géntermékek elnevezése
VPg- genomhoz kötött fehérje, P1- első fehérje, HC-Pro- segítő komponens-proteáz, P3- harmadik fehérje, 6K1- az első 6 kDa-os fehérje, CI- henger alakúzárványfehérje, 6K2- a második 6 kDa-os fehérje, NIa (VPg-Pro)- kis nukleáriszárványfehérje, NIb- nagy nukleáris zárványfehérje, CP- köpenyfehérje, polyA-poliadenilált régió.
A potyvírusok által termelt fehérjék nagy részének funkciója többnyire
ismert, de teljesen még nem tisztázott. A fehérjék többsége általában többféle
folyamatban is részt vesz. Egyes fehérjék szerepére csak más, ismert funkciójú
fehérjékhez való hasonlatosság alapján következtetnek. A P1 proteináz a
P1/HC-Pro hasítást végzi, rendelkezik nukleinsav kötő tulajdonsággal, aminek a
sejtről sejtre való terjedésben lehet szerepe, illetve részt vehet a
tünetfejlődésben és szinergista kölcsönhatásokban (Verchot és mtsai, 1991,
Soumounon és Laliberté, 1994, Pruss és mtsai, 1997). A HC-Pro proteináz a
HC-Pro/P3 hasítást végzi, részt vesz a levéltetűvel történő átvitelben, a sejtről-
sejtre- és a hosszú távú terjedésben, de szerepe lehet még a virulencia és a
tünetek erősségében (Carrington és mtsai, 1989, Maia és mtsai, 1996, Rojas és
mtsai, 1997, Kasschau és mtsai, 1997). A P3 feltételezett szerepe a
tünetfejlődésben lehet, valamint a vírusreplikációban (Rodrigez-Cerezo és
mtasi, 1993, Riechmann és mtsai, 1995), a 6K1 szerepe nem ismert, a 6K2 a
vírusreplikációban vesz részt (Riechmann és mtsai, 1992). A CI helikáz
aktivitással rendelkezik és a sejtről-sejtre történő terjedésben vesz részt
(Rodriguez-Cerezo és mtsai, 1997, Carrington és mtsai, 1998). Minden
potyvírusnál megtalálható jellegzetes szélkerék alakú zárványok formájában
P1 Hc-Pro P3
6K1
CI
6K2
NIa NIb CP
poly A
VPg

16
(Brakke és mtsai, 1987). Az NIa kis sejtmagi zárványfehérje proteáz
aktivitással rendelkezik (Carrington és Dougherty, 1987), részt vesz a hosszú
távú mozgásban, N-terminális doménje a genomhoz kötött protein (VPg),
amely a replikáció során kötődik a vírus RNS-hez (Murphy és mtsai, 1990). A
NIb RNS függő RNS polimeráz (Shukla és mtsai, 1994), a CP a vírus RNS
becsomagolásában, a vektorokkal való terjedésben (DAG motívum), a hosszú
távú mozgásban és a genom amplifikációban vesz részt (Shukla és mtsai, 1994,
Haldeman-Cahill és mtsai, 1998, López-Moya és Pirone, 1998). Az 5’-NCR
(nem kódoló régió) részt vesz az enkapszidációban, valószínűsíthető, hogy a
transzlációban és a replikációban, illetve szerepe lehet a fertőzőképesség
alakulásában is (Shukla és mtsai, 1994, Xu és Shaw, 1998, Palkovics és mtsai,
1998). A 3’-NCR pedig a vírusreplikációban vesz részt (Haldeman-Cahill és
mtsai, 1998).
A potyvírusok mechanikai sérüléseken keresztül jutnak be a gazdaszervezet
sejtjeibe, amelyeket fizikai behatások vagy a vírus vektorai okoznak. A
potyvírusok replikációs ciklusa a növényi sejtbe való bekerülést követően a
vírus RNS kicsomagolódásával, majd a transzlációt követő poliprotein
hasítással folytatódik. A genomreplikáció a negatív szál intermedieren keresztül
történik. A vírus a gazda által termelt energiát és anyagokat használja
fennmaradásához és replikációjához, önmaga csak a genomi RNS-ben tárolt
információt szolgáltatja.
A szisztemikus és lokális vírustünetek eltérő jellege alapján több PVY törzs
különböztethető meg. Magyarországon túlnyomó többségben a PVYNTN törzs
fordul elő (Wolf és Horváth, 2000). Az általános PVYO törzs, az egész világon
elterjedt (Ellis és mtsai, 1997) és többféle kórképet okoz. Gyűrődéses,
nekrotikus szimptómákat eredményez burgonyán valamint levélszáradást,
fodrosodást, levélhullást, dohányon pedig foltokat, szisztemikus

17
márványozottságot (Smith és Dennis, 1940, Horváth, 1966). A PVYN törzs
szintén előfordul az egész világon. Érnekrózist okoz dohányon, paprikán és
paradicsomon, valamint enyhe foltosodást hoz létre a legtöbb termesztett
burgonya esetében. Magyarországi előfordulását Szirmai (1958) írta le először.
A PVYC törzs előfordulása Ausztráliában, Európában, Indiában és Dél-
Afrikában ismert. Burgonyán hiperszenzitív reakciókat vált ki, nekrotikus
léziókat foltokkal a levélen és a száron (de Bokx és Huttinga, 1981). A
dohányon okozott tünetek megegyeznek a PVYO törzs által okozottakkal. A
PVYAn (anomale törzs) az N törzshöz hasonlóan érnekrózist okoz, de
szerológiailag az O és az N törzsek között helyezkedik el. A PVYNTN (new
tuber necrosis) törzs, mely a burgonya gumó nekrotikus gyűrűsfoltosság nevet
kapta, Európában és a világ számos más országában is elterjedt. A burgonya
levelén nekrotikus léziókat, a gumón és a bogyón nekrotikus gyűrűsfoltosságot
okoz, és rezisztencia-áttörő tulajdonsága miatt jelentős. A gumók
tárolhatatlanná válnak, sérülékenyekké és eladhatatlanokká. A megemlített
fontosabb törzsek mellett még előfordulnak PVYNN, PVYMM és PVYMN törzsek
is, melyek csoportosítása az általuk okozott tünetek alapján történt (Gooding és
Tolin, 1973, Sudarsono és mtsai, 1993, Chachulska, 1998).
A magyarországi flórából izolált burgonya Y vírus (PVY-H), mely a
PVYNTN törzsébe tartozik, 9703 nukleotid hosszúságú genomi RNS-sel
rendelkezik és egy poli(A) véggel (Thole és mtsai, 1993). A PVY-H nukleotid
szekvenciája egy nagy olvasási keretet tartalmaz, amely egy 3061 aminosav-
hosszú poliprotein prekurzort kódol, valamint egy 189 nukleotidból álló 5' nem-
kódoló régióval és egy 330 nukleotid-hosszú 3' nem transzlálódó régióval
rendelkezik. A PVY törzsek általában nagyobb mértékű szekvencia
heterogenitással rendelkeznek, mint más potyvírus törzsek, amelyek nukleotid

18
szekvencia azonossága 95 %-nál magasabb. (Robaglia és mtsai, 1989, Shukla
és mtsai, 1991).
2.3. PATOGÉNTŐL SZÁRMAZTATOTT REZISZTENCIA
A Patogéntől Származtatott Rezisztencia (PDR) fogalma 1985-ből
Sanfordtól és Johnsontól ered. E megfogalmazás szerint a gazdanövényben
expresszálódó vírus génjének, génszakaszának RNS transzkriptuma, illetve
génterméke a víruseredetű komponensek természetes egyensúlyát
megváltoztathatja, így a vírus replikáció, valamint a vírus növényen belüli
terjedése megakadályozható, illetve megzavarható, ami a növény enyhített vagy
elmaradó fertőzését eredményezi.
2.3.1. Klasszikus keresztvédettség
A növénykórtanban jól ismert a keresztvédettségi reakció, amelyet a
gyakorlati mezőgazdaság számos esetben alkalmaz sikerrel (visszaszorítva a
vad típusú vírus által okozott betegségtüneteket illetve a vírus replikációját). A
gyengített vírustörzzsel kialakított klasszikus keresztvédettség jelenségét
először McKinney írta le 1929-ben. TMV (tobacco mosaic virus) gyengített
törzsével történő fertőzés esetén azt tapasztalta, hogy a későbbi vírustámadásnál
a növény a vírus különböző vonalaival szemben rezisztens volt. Üvegházi
vizsgálatokkal kimutatták, hogy a fertőzés után a felülfertőző vírus csak kis
mennyiségben van jelen a növényben, esetenként ki sem mutatható. A
rezisztencia nem jelenik meg minden esetben, például "A" vírus nyújthat
védelmet "B"-vel szemben, míg "B" már nem "A"-val szemben (Palukaitis és

19
Zaitlin, 1984). Általánosságban elmondható, hogy főképpen rokon törzsek
között figyelhető meg a jelenség kialakulása, ami szekvencia homológiára
vezethető vissza.
A jelenség felismerését követően többféle hipotézis született a klasszikus
keresztvédettségi reakció működésének magyarázatára. Zaitlin 1976-ban
felvázolta az RNS : RNS interakciók egyik lehetséges módját. Azt feltételezte,
hogy a keresztvédettség molekulák közötti hibridizáció eredménye, a gyengített
vírus RNS és a fertőző vírus RNS között. De Zoeten és Fulton szerint (1975)
egy ún. második becsomagolódása történik a fertőző vírus RNS-nek, a
gyengített vírustól származó szabad köpenyfehérjébe. 1982-ben Sherwood és
Fulton azt feltételezte, hogy a fertőző vírus képtelen kicsomagolódni, ha
gyengített vírussal szisztemikusan fertőzött növényt támad. Zinnen és Fulton
(1986) keresztvédettségi kísérletet végeztek SHMV (sunn hemp mosaic virus)
és TMV felhasználásával. Azt tapasztalták, hogy a védettség kialakítása
céljából gyengített SHMV-vel fertőzött növényt, ha TMV RNS-el fertőznek és
az SHMV CP-be becsomagolódik, 5-27-szer kevésbé lesz fertőzőképes, mint
amikor TMV CP-be csomagolódik be az RNS. Azt a következtetést vonták le,
hogy a szabad, homológ CP képes csökkenteni a fertőzőképességet, de
feltételezték, hogy egyéb, nem ismert tényezők is szerepet játszhatnak a
folyamatban. 1981-ben Sarkar és Smitamana, valamint 1989-ben Gerber és
Sarkar olyan mutáns, CP nélküli TMV vonalat állítottak elő, amely szintén
képes volt keresztvédettséget kiváltani. Ezek alapján arra következtettek, hogy
nagyobb szerepük lehet a rezisztencia kialakításában az addig még nem ismert
tényezőknek, mint magának a CP-nek. Más vélemények szerint a gazdasejtben
valamiféle verseny alakul ki a védő vírus és a fertőző vírus között a
replikációhoz szükséges valamely komponensért. Ez lehet például maga a
replikáz gén. Horikoshi és mtsai (1987) leírták, hogy a BMV (brome mosaic

20
virus) CP blokkolta a replikáz kötőhelyét, ezáltal megzavarta az RNS szintézist.
Azt feltételezték, hogy a CP által szabályozott vírusreplikáció képezi a
keresztvédettség hatásmechanizmusának alapját.
Az módszer azonban nem terjedt el, mert ezek a gyengített vírustörzsek
más gazdanövényeken súlyos betegséget, esetleg teljes pusztulást okoztak. Az
attenuált törzsek felhasználása magában hordozza azt a lehetőséget is, hogy
mutáció során az eredetileg gyengített vírustörzs patogenitása megváltozik.
További kockázati tényezőt jelenthet a szinergisztikus hatás, mely valamely
más vírussal való együttes fertőzésekor léphet fel. Ekkor a két vírus egyidejű
jelenléte ugyanazon növényben fokozottabb kártételt eredményez, mint a kettő
külön-külön.
A molekuláris biológiai módszerek fejlődésével azonban lehetőség nyílt a
növénykórtanban jól ismert keresztvédettségi reakció kialakítására,
hatékonyabb és biztonságosabb módon, a gyengített vírus által hordozott
kockázati tényezők nélkül..
2.3.2. Molekuláris módszerekkel kiváltott ellenállóképesség
A molekuláris biológia módszereinek fejlődésével lehetőség nyílt arra,
hogy a vírus eredetű gének növényi genomba építésével és azok növényen
belüli megnyilvánításával az előzőekben ismertetetthez hasonló
keresztvédettségi reakciót alakítsanak ki. Széles körben és sikeresen használt a
vírus köpenyfehérje génje, melyet elsőként alkalmaztak a klasszikus
keresztvédettség vizsgálatainak eredményei alapján. Transzgénikus növények
előállításához a köpenyfehérje gén mellett számos más vírusgén is
felhasználható: pl. a replikázkomplex komponenseinek génjei, szatellit RNS-ek,

21
defektív interferáló elemek, ribozimok, antiszensz RNS-ek és mozgási fehérjék
génjei (Wilson, 1993, Lindbo és mtsai, 1993b, Baulcombe, 1996, Tacke és
mtsai, 1996, Ares és mtsai, 1998).
2.3.2.1. Köpenyfehérje (CP) jellemzése
A köpenyfehérje a legjobban jellemzett fehérje a potyvírusoknál.
Elsődleges feladata a vírus RNS becsomagolása, de ezen túl részt vesz a
levéltetvekkel történő terjedésben (Pirone, 1991, Salomon és Bernardi, 1995), a
tünetek kialakításában és a sejtről-sejtre történő terjedésben (Dolja és mtsai,
1995, López-Moya és Pirone, 1998, Varrelman és Maiss, 2000), de kimutatták a
CP replikációban betöltött szerepét is (Mahayan és mtsai, 1996).
A CP egy erősen konzervált központi magból és a változékonyabb N- és C-
végekből áll. A virion felületén helyezkednek el a CP N- és C-terminális részei
(Ward és Shukla, 1991), melyek nem szükségesek a víruspartikulumok
összeszerelődéséhez, sem a mechanikai inokulációval történő fertőzéshez
(Shukla és mtsai, 1994). Az N-terminális szakasz, mely mind hosszban, mind
szekvenciáiban változékony, részt vesz gazda-vektor-vírus kölcsönhatásokban,
alkalmas víruscsaládon belüli elkülönítésre és izolátumok megkülönböztetésére,
azonban nem játszik szerepet a sejtről-sejtre történő terjedésben és a
plazmodezmákkal kialakított kapcsolatban (Rojas és mtsai, 1997) (5. ábra).

22
5. ábra
A PVY CP harmadlagos struktúrájának sematikus vázlata
(Shukla és Ward, 1989)
Az N-terminális végen elhelyezkedő DAG triplet a levéltetvek általi
átvitelben játszik fontos szerepet (Salomon és Raccah, 1990). A CP a segítő
komponenssel kapcsolatba lépve egyes feltevések szerint a levéltetvek
szájszervéhez kapcsolódik (Pirone és Blanc, 1996). A C- és N-terminális régió
felelős a növényen belüli hosszú távú mozgásért (Dolja és mtsai, 1995). A vírus
nukleoprotein komplexet alkotva jut át a plazmodezmákon (Rodriguez-Cerezo
és mtsai, 1997). Az enkapszidációhoz viszont nem feltétlenül szükségesek,
eltávolításuk nem befolyásolja a vírus fertőzőképességét (Shukla és mtsai,
1994). A potyvírusok képesek másik, de a nemzetségbe tartozó vírus
köpenyfehérje alegységeit is felhasználni a burok felépítéséhez, így
levéltetvekkel át nem vihető vírus átvihetővé válik. Ez a heteroenkapszidáció
jelensége.

23
2.3.2.2. A köpenyfehérje gén által közvetített rezisztencia (coat protein
mediated resistance -CPMR)
A második vírustörzzsel történő fertőzéskor fellépő rezisztenciát a növényi
sejtekben már megtalálható vírus eredetű köpenyfehérje jelenlétével is
magyarázták (de Zoeten és Fulton, 1975). A biotechnológiai módszerek
fejlődésével felismerték a lehetőséget, hogy a köpenyfehérje génnel történő
transzformáció segítségével a keresztvédettséghez hasonló eredmények érhetők
el. A gént megfelelő vektorba építve és növényi genomba integrálva a
köpenyfehérje növényen belüli termelődését lehet kiváltani. Az így létrehozott
rezisztenciát nevezzük köpenyfehérje által közvetített rezisztenciának. A
védettség mértéke függ az adott vírustól, az inokulum koncentrációjától és a
transzformált növényi vonaltól. A rezisztencia szintje azonban változó: a
szimptómák kifejlődése időben késhet, a megjelenő tünetek lehetnek
enyhébbek, előfordulhat alacsonyabb vírusszám, létrejöhet felépülési reakció
(„recovery”: amikor a növény elsődlegesen fertőződik, azonban a később
megjelenő fiatal levelek már tünet- és vírusmentesek), valamint kialakulhat
immunitás. A védettség esetenként fennállhat magasabb inokulum-
koncentrációval szemben is.
Az első kísérletet köpenyfehérje közvetítette rezisztencia kialakítására
Bevan és munkatársai 1985-ben végezték. Dohány mozaik vírus (tobacco
mosaic tobamovirus -TMV) köpenyfehérje (coat protein -CP) gént építettek be
dohánynövényekbe Agrobacterium tumefaciens segítségével. A felnevelt
növények leveleiből 0.001 %-ban mutatták ki a CP-t az összes oldható
fehérjéhez viszonyítva, azonban TMV fertőzéskor a transzgénikus növények a
kontrollokhoz hasonlóan fogékonynak bizonyultak.

24
Mintegy 18 évvel ezelőtt Powell-Abel és munkatársai (1986) szintén TMV
köpenyfehérje gént építettek be dohány és paradicsom növényekbe, amelyek
ezáltal nagyfokú ellenállóképességet mutattak a felülfertőző vírussal szemben.
A TMV CP génjét tartalmazó konstrukcióval transzformálták a növényeket,
mely konstrukció a 35 S promótert és poly (A) régiót is tartalmazta. A
transzgénikus növények leveleiből származó fehérjekivonatban immunoblot
segítségével 0.1 %-os arányban mutatták ki a vírus köpenyfehérjét. TMV-vel
történő mesterséges fertőzés során a CP-t szintetizáló növényeken késéssel
jelentkeztek a tünetek, illetve egy részük tünetmentes is maradt. A
transzformánsokon megfigyelt rezisztenciát a hasadó utódpopulációkon is ki
lehetett mutatni.
Ez volt az első sikeres tanulmány, amely bebizonyította, hogy
vírusrezisztencia kialakításának céljából a genetikai transzformációt
eredményesen lehet alkalmazni. Ezáltal egy új, korszerű génsebészeti eljárás
felhasználásával vált lehetővé a vírus-ellenállóság kialakítása. Ma már több
mint száz vírus-gazdanövény kapcsolatban alkalmazták sikeresen az ún.
„köpenyfehérje gén által közvetített ellenállóság” jelenségét. Az utóbbi években
az így előállított transzgénikus növények közül számos köztermesztésbe is
került.
2.3.2.3. Köpenyfehérje által közvetített keresztvédettség
1986 óta CP által kiváltott védettséget számos esetben hoztak létre a világ
különböző laboratóriumaiban, mintegy 16 vírusnemzetség egyes tagjaival
szemben és több gazdaságilag fontos növényfaj esetében. Az 2-es számú
melléklet tartalmazza az e célra már felhasznált vírusnemzetségek és

25
növényfajok összefoglaló táblázatát, a 1-es számú mellékletben pedig ugyanezt
tárgyaljuk kissé részletesebben, az egyes vírusnemzetségek szerint csoportosítva.
2.3.2.3.1. Potyvirus nemzetség
Különböző laboratóriumokban számos sikeres kísérletet végeztek a
Potyvirus nemzetség több tagjával, így a PVY felhasználásával is.
Cassidy és Nelson (1995) például a földimogyoró csíkosság vírus (peanut
stripe potyvirus –PStV) CP gént tartalmazó konstrukciókkal transzformáltak N.
benthamiana növényeket. Létrehoztak ATG transzlációs start kóddal
rendelkező és e nélküli, teljes hosszúságú konstrukciókat, valamint ATG-vel
rendelkező, de csonkított CP-t tartalmazó vektorokat. A két -teljes gént
tartalmazó- „ATG+” konstrukciókkal transzformált vonalaknál mechanikai
fertőzés esetén nem alakultak ki vírustünetek és vírusakkumuláció sem volt. A
többi vonalnál csupán a tünetfejlődés késését tapasztalták. A később fejlődő
szisztemikus, fiatal levelek tünetmentesek voltak és a CP-t sem lehetett belőlük
kimutatni. További vizsgálatokkal bebizonyították, hogy a felépülési „recovery”
jelenséget mutató növények tünet nélküli levelei rezisztensek az újabb PStV
fertőzésekkel szemben, azonban TEV fertőzéssel szemben nem. A PStV-re
immunis transzgénikus növények ellenállóképessége nem fejeződött ki más
poty- vagy tobamovírusokkal történő fertőzés esetén. A vizsgálatok nem
mutattak ki korrelációt a CP akkumulációs szint és a rezisztencia mértéke
között.
Dinant és mtsai (1993) saláta mozaik vírus (lettuce mosaic potyvirus –
LMV) CP génnel transzformáltak N. tabacum cv. Xanthi dohánynövényeket,
majd ezeket PVY-nal fertőzték. A 34 vizsgált növényi vonalból 17 esetben

26
mutattak ki CP akkumulációt. Ezek a vonalak rezisztensek voltak PVYN
fertőzéssel szemben, holott PVY és az LMV CP-k közötti aminosavhomológia
csupán 66 %. A második generációból 5 vonalat vizsgálva azt tapasztalták,
hogy a növények ellenállóak voltak a PVYN és még négy másik PVY törzzsel
szemben is. Két vonalban teljes rezisztenciát tapasztaltak, két másik vonalban
csupán a tünetek kialakulásának késését, egy vonalban pedig a kontrollokhoz
hasonló erősségű tüneteket figyeltek meg, melyek késve, de megjelentek.
Murry és mtsai (1993) kukorica törpe mozaik vírus (maize dwarf mosaic
potyvirus –MDMV) 'B' törzsének CP génjét felhasználva állítottak elő
transzgénikus kukoricanövényeket. A vonalak mindegyike expresszálta a CP
gént. A növényeket MDMV 'A' és 'B' törzsekkel fertőzték, valamint kukorica
klorotikus foltosság vírussal (maize chlorotic mottle machlomovirus –MCMV)
együtt, keverten. A transzgénikus növényeket egyik vírus sem tudta
megfertőzni. A replikácó és a vírusterjedés gátlása a heterológ fertőzésnél nem
volt teljes, a transzgén jelenléte mégis elősegítette a növények életben
maradását.
Lindbo és Dougherty (1992b) nem-transzlálódó, szensz és antiszensz
dohány karcolatos vírus (tobacco etch virus –TEV) CP génnel transzformáltak
dohánynövényeket. A 7 vizsgált, antiszensz gént hordozó vonalból 1 esetben
tapasztaltak tünetgyengülést, míg a tíz szensz vonalak mindegyike tünetmentes
volt. Mechanikai és levéltetvek általi inokulációt is alkalmaztak. A védettség
TEV specifikus volt, mely elsősorban a vírus replikáció csökkentésében
nyilvánult meg, és független volt az inokulum koncentrációjától és a növény
méretétől. A rezisztencia RNS-függőnek bizonyult, mivel fehérjetermék
hiányában is megnyilvánult.
Ravelonandro és mtsai (1993) PPV CP génnel transzformáltak N. tabacum
cv. Xanthi, N. benthamiana és N. clevelandii növényeket. A N. benthamiana

27
vonalakban ellenállóképesség kialakulását tapasztalták PPV fertőzés esetén,
míg a N. tabacum cv. Xanthi és a N. clevelandii vonalakban csupán részleges
védettséget figyeltek meg PPV és PVY inokulációkat követően. 20 µg/ml PVY-
nal történő fertőzés esetében a vonalak 90-100 %-a megfertőződött, ami a
rezisztencia inokulum-koncentrációtól való függőségét jelzi. Egyes N.
benthamiana növényeknél a hosszú távú vírusmozgás blokkolását és a
felépülési jelenséget figyelték meg, ez azonban szintén függött az alkalmazott
víruskoncentrációtól.
Stark és Beachy (1989) szója mozaik vírus (soybean mosaic potyvirus –
SMV) 'N' törzs CP génjét használták fel transzgénikus dohánynövények
előállításához. A növények ellenállóak voltak PVY-nal és TEV-vel szemben,
míg dohány mozaik vírus (tobacco mosaic virus –TMV) 'U1' fertőzés esetén
fogékonyként viselkedtek.
Dohány érfoltosság vírussal szemben (tobacco vein mottling potyvirus –
TVMV) Murphy és mtsai (1990) hoztak létre rezisztens transzgénikus
dohányvonalakat, melyek nemcsak TVMV-vel szemben voltak ellenállóak,
hanem TEV fertőzéssel szemben is rendelkeztek rezisztenciával.
PVY 'O' törzsének CP génjével végeztek transzformációt Lawson és mtsai
(1990) és azt tapasztalták, hogy a transzgénikus Russet Burbank
burgonyanövények védettek voltak PVY 'O' fertőzéssel szemben, míg PVX-szel
szemben fogékonynak mutatkoztak.
Scorza és mtsai (1991) papaya gyűrűsfoltosság vírus (papaya ringspot
potyvirus –PRSV) CP génnel transzgénikus szilvákat fertőztek PPV-vel
(átoltással) és két éven keresztül vizsgálták a reakciókat. A legtöbb
transzgénikus növény megfertőződött, viszont a tünetek a kontrollokhoz képest
enyhébbek voltak, illetve egy esetben a védettség megjelenését is tapasztalták.

28
Ling és mtsai (1991) szintén PRSV CP gént expresszáltattak
dohánynövényekben. TEV, PVY és paprika foltosság vírus (pepper mottle virus
–PepMoV) fertőzést követően a tünetek gyengült és késő kifejlődését figyelték
meg. Az ellenállóképesség TEV és PepMoV esetében hatékonyabbnak
bizonyult, mint PVY-nal szemben.
Da Camara Machado és mtsai (1992) Prunus armeniaca sejtkultúrát
transzformáltak szilva himlő vírus (plum pox potyvirus –PPV) CP génjét
hordozó konstrukcióval. A sikeres kajszibarack regeneráció fejletlen
embriókból történt. Ez volt az első tanulmány, mely eredményes gyümölcsfa-
transzformációról számolt be.
Kollár és mtsai (1993) szintén PVY köpenyfehérje génnel transzformáltak
N. tabacum növényeket. A CP-t expresszáló vonalak rezisztensek voltak PVY
fertőzéssel szemben azonban a vizsgálatok során kimutatták, hogy a
rezisztencia nem függ a CP jelenlététől. Hasonló eredményeket kaptak van der
Vlugt és mtsai (1992) dohánynövények és PVY 'N' izolátum esetén.
Fang és Grumet (1993) cukkíni sárga mozaik vírus (zucchini yellow
mosaic potyvirus –ZYMV) CP-t tartalmazó konstrukció felhasználásával
hoztak létre ZYMV fertőzéssel szemben ellenállóságot sárgadinnyében.
Scorza és mtsai (1994) szilva hipokotil szeleteket transzformáltak PPV CP
génnel. 1800 szeletből 22 transzformáns klónt kaptak, melyek közül 5 termelte
a CP-t. A mRNS szint a kimutathatatlantól a nagy mennyiségig terjedt és a CP
mennyiség korrelált a mRNS szinttel. A termelődött köpenyfehérje
mennyiségét a beépülési kópiaszámmal és az integráció helyével hozták
összefüggésbe.
1994 -ben Malnoë és mtsai számoltak be sikeres szántóföldi kísérletekről
PVY CP génnel transzgénikus Bintje burgonyafajta esetében. Azt tapasztalták,

29
hogy a rezisztencia levéltetvekkel történő természetes fertőzés esetén is
megmaradt, sőt a gumóval történő vírusterjedés is lelassult vagy meggátlódott.
Palkovics és munkatársai (1995) az SK-68 jelű PPV izolátum
köpenyfehérje génjét építették be N. benthamiana növényekbe. Két vonal
esetében a „recovery” jelenséget figyelték meg, míg a vonalak nagy része
rezisztens volt a fertőző PPV vírussal szemben..
Regner és mtsai (1992) is hasonló eredményekről számoltak be, azonban N.
clevelandii növények esetében is megfigyelték a fertőzést áttörő, új,
tünetmentes levelek kialakulását. A rezisztencia szintje és a CP expresszió
között ugyanakkor fordított korrelációt figyeltek meg.
Smith és mtsai (1995) PVY CP-vel transzformáltak burgonyanövényeket,
melyek vizsgálata során szintén fordított korrelációt figyeltek meg a
köpenyfehérje akkumuláció és a rezisztencia erőssége között. A magas fokú
ellenállóképességet kifejező vonalak több transzgén kópiát tartalmaztak és
alacsony alapállapotú RNS szinttel rendelkeztek. Ez a megfigyelés általánosan
is jellemző a potyvírus eredetű CP gének által kiváltott ellenállóképesség
molekuláris hátterére.
2.3.2.4. A köpenyfehérje gén által indukált rezisztencia lehetséges
hatásmechanizmusai
A vírusszekvenciákkal transzformált növények ellenállóságát előidézhetik
a fehérjék vagy a víruseredetű nukleinsavak. A proteinek által közvetített
rezisztencia általában széles spektrumú, azaz többféle vírussal szemben nyújthat
védelmet, azonban gyengébb hatékonyságú és vírus nukleinsavval történő
fertőzés esetén áttörhető. Az RNS-ek által közvetített ellenállóság magas
inokulum koncentrációval szemben is hatékony, a növény korától független

30
(fiatal növény esetében is működőképes). Az utóbbi nemcsak virionnal, hanem
nukleinsavval történő fertőzést követően is megmarad és protoplaszt-szinten is
fennállhat, viszont sokkal inkább vírus specifikus. A csonkított vagy kiméra
CP-k alkalmazása bizonyos esetekben bővítheti a rezisztencia hatásspektrumát
illetve fokozhatja hatékonyságát. Nem mutattak ki minden esetben egyértelmű
korrelációt a CP akkumuláció és a rezisztencia szintje között, sőt, esetenként
CP-t akkumuláló vonalak a nem-transzformáns vonalakhoz hasonlóan
érzékenyek voltak a fertőzéssel szemben. A rezisztencia erőssége függ az adott
vírustól és a transzformáns növényi vonaltól. A rezisztencia mértéke változó,
előfordulhat csupán a tünetek erősségének csökkenése vagy időben későbbi
kifejlődése, de megfigyelhető a tünetmentesség és az immunitás is. Előfordult,
hogy a növények először megfertőződtek és mutatták a tipikus tüneteket, majd
idővel teljesen tünetmentessé váltak (Hammond és Kamo, 1993., Lindbo és
Dougherty, 1992a).
2.3.2.4.1. A köpenyfehérje által közvetített rezisztencia hatásmechanizmusai
A CP által közvetített rezisztencia esetében egyenes arányosság áll fenn az
ellenállóság mértéke és a proteinakkumuláció szintje között. A védettség intakt
virionokkal történő fertőzést követően nyilvánul meg, vírus RNS-t használva
inokulumként a rezisztencia áttörhető. Működésének magyarázatára számos
hipotézis született, mivel a proteinek által közvetített rezisztencia valószínűleg
többféle mechanizmus által fejti ki hatását a gazda-vírus-transzgén
kombinációtól függően. Előfordulhat, hogy a fertőző vírus RNS-ének egy ún.
második becsomagolódása, enkapszidációja történik a transzgénről képződő
CP-be, így gátolva annak terjedését. Lehetséges az is, hogy már a vírus
dekapszidációja gátolt. Voltak olyan feltételezések is, hogy a köpenyfehérje a

31
vírus RNS replikációját befolyásolhatja (Baulcombe és English, 1996),
valamint gátolhatja a vírus RNS kötődését a riboszómákhoz (Clark és mtsai,
1995). A legtöbb köpenyfehérje által indukált rezisztencia esetében a fertőzést
követően a szisztemikus tünetek nem fejlődnek ki, vagy kialakulnak, de
késéssel. Ez a jelenség a vírus sejtről-sejtre történő terjedésének, a
szállítószövet-rendszerbe és a nem-inokulált levelekbe való bejutásának
gátlásából adódhat, vagy a fertőzés kezdetének megakadályozásából (Beachy,
1990, Wisniewski és mtsai, 1990). Egyes vírusoknál, mint pl. a TMV,
kísérletekkel alátámasztva mutatták ki a fehérjetermék közvetlen szerepét a
rezisztencia kiváltásában (Powell-Abel, 1986). Powell-Abel és mtsai (1990)
AUG transzlációs start kodon nélküli TMV CP-t tartalmazó konstrukcióval
transzformáltak növényeket, melyek fogékonynak bizonyultak TMV fertőzéssel
szemben, bizonyítva, hogy ez esetben nem a mRNS a rezisztencia kiváltója,
hanem a fehérjetermék. Van Dun és mtsai (1988) csonkított AMV CP-t
állítottak elő, amely nem volt alkalmas a rezisztencia kiváltására. A
köpenyfehérjét expresszáló növényekből nyert protoplasztokkal végzett
kísérletek alapján megállapították, hogy a transzgén által termelt CP a fertőző
vírus kicsomagolódásába avatkozik be, így replikációs ciklusának kezdeti
lépését gátolja. Ennek módja lehet a vírus szét- és összeszerelődési
egyensúlyának eltolása az összeépülés irányába, de előfordulhat, hogy a sejten
belüli vírus partikulum szétszerelődéshez szükséges speciális helyek vagy
receptorok lekötöttségében bekövetkezett változások okozzák a
kicsomagolódás-gátlást. A transzgén eredetű CP alegységek vírusszerű
partikulumokat (VLP) képezve elfoglalják ezeket a helyeket, így
megakadályozzák a fertőzést (Lomonosoff, 1995). Éppen ezért a CP által
indukált rezisztencia függ az elsődlegesen fertőzött epidermális sejtekben
megnyilvánuló CP produkciótól (Reimann-Philip és Beachy, 1993). Ezt olyan

32
kísérletekkel bizonyították, amelyekben a levél mezofillumsejtekben
expresszáltatták a CP-t specifikus promóterrel, rezisztencia kialakulása nélkül.
A CP által indukált rezisztencia kialakításához általában vad típusú vírus
köpenyfehérje génjét használják, azonban Bendahmane és mtsai (1997) olyan
mutáns TMV CP konstrukciókkal transzformáltak növényeket, melyek
köpenyfehérje alegységei közötti kölcsönhatások kísérletenként eltérőek voltak.
TMV fertőzés esetén azok a növényi vonalak bizonyultak rezisztensnek,
melyek transzgénjéről expresszálódó mutáns CP alegységek közötti
kölcsönhatás erős volt, mivel képesek voltak vírusszerű partikulumokat
létrehozni, ezáltal elfoglalni a fertőző vírus szétszerelődéséhez szükséges
receptorhelyeket. Az így kialakult rezisztencia magasabb fokú volt, mint a vad
típusú vírus CP-k által kiváltott. Ezek az eredmények arra utalnak, hogy a
köpenyfehérje által indukált rezisztencia kialakításában fontos szerepet
játszanak a transzgén eredetű- és a fertőző vírus eredetű köpenyfehérje
alegységek közötti kölcsönhatások.
2.3.2.4.2. Az RNS-ek által közvetített rezisztencia hatásmechanizmusai, a
géncsendesítés jelensége
A köpenyfehérje által közvetített rezisztencia tanulmányozásában számos
olyan példa található, amelyekben a rezisztencia a transzgénről származó
fehérjetermék hiányában is kialakul. Ilyenkor a transzgénikus növényekben a
védettséget maga a transzgén vagy az erről átíródott transzkriptum váltja ki. A
jelenséget megfigyelték pl. a TSWV köpenyfehérje génnel való
transzformációkor De Haan és mtsai (1992), PVY esetében Van der Vlugt és
mtsai (1992) és TEV-nél Lindbo és Dougherty (1992b). Az RNS által

33
közvetített rezisztencia, amely a géncsendesítés (gene silencing) mechanizmusa
által nyilvánul meg, jellemzően alacsony vagy nem detektálható transzgén
expresszióval jár együtt. Mechanizmusának kutatását előremozdította a
„recovery” jelenség felfedezése és vizsgálata (Lindbo és Dougherty, 1992a,
Lindbo és mtsai, 1993b). TEV-vel inokulált transzgénikus növények kezdetben
mutatták a fertőzés tüneteit, majd 3-5 héttel később a fiatal, kifejlődő levelek
tünet- és vírusmentesek voltak. A felépült növények a későbbi TEV fertőzéssel
szemben is ellenállóak voltak. A növényeknek számos módszerük van a
vírusfertőzés elkerülésére, ezek egyike a géncsendesítés, amikor a vírus RNS
egy szekvenciaspecifikus mechanizmus által degradálódik (Carrington és mtsai,
2001). Az antiszensz konstrukciókkal kiváltott rezisztencia gyakran nagyobb
hatékonyságú, mint ugyanaz a szekvencia szensz orientációban (Smith és mtsai,
1995).
Az RNS által közvetített rezisztencia egyik fajtája a homológia-függő
rezisztencia, mivel kialakulása a transzgén és a célszekvencia (vírus RNS)
közötti homológiától függ (Sonoda és mtsai, 1999, Mäki-Valkama és mtsai,
2000). A jelenség működésének magyarázatára egy kvantitatív és egy kvalitatív
modellt állítottak fel. A kvantitatív modell szerint ha a transzgén eredetű RNS
szint meghalad egy küszöbértéket, aktiválódik egy citoplazmatikus RNS
degradáló mechanizmus, mely lebontja az endogén és az ezzel homológ exogén
vírus RNS-eket. Ezzel a modellel magyarázták a felépülési jelenséget is. A
küszöbérték átlépéséhez azonban nagy transzkripciós aktivitás vagy nagyobb
beépülési kópiaszám szükséges (Lindbo és mtsai, 1993a, Smith és mtsai, 1994,
Goodwin és mtsai, 1996). A kvalitatív jellegű modell alapeleme a normál
poliadenilált traszkriptumoktól eltérő, un. aberráns RNS-ek képződése, melyek
egy szekvenciaspecifikus RNS degradáló mechanizmus által lebontódnak
(Mueller és mtsai, 1995, English és mtsai, 1996). A géncsendesítésnek két

34
fajtája van: egyik a transzkripcionális géncsendesítés, mely a sejtmagban
nyilvánul meg. Ekkor a gének nem íródnak át DNS metiláció vagy promóter
inaktiváció következtében. A másik a poszttranszkripcionális géncsendesítés,
mely a citoplazmában történik. Ez esetben a gének átíródnak, de a
transzkriptumok már degradálódnak. A poszttranszkripcionális
géncsendesítésnek (hatástalanításnak) három szakasza különíthető el:
kiváltódás, terjedés és fenntartás (Voinnet és mtsai, 1998, Palauqui és
Balzergue, 1999) és három fő tulajdonsága van: adaptív, mobilis és specifikus
(Voinnet, 2001). A mechanizmus aktiválódhat transzgén vagy vírus eredetű,
nagy mennyiségű duplaszálú RNS (dsRNS) molekulák által (mivel egészséges,
nem transzgénikus növényben dsRNS nincs). A mechanizmus eredményezheti
a homológ RNS-ek degradációját a citoplazmában és a homológ nukleáris
DNS-ek metilációját a sejtmagban (Matzke és mtsai, 2001). A dsRNS-ek az
RNS vírusok replikációjának köztitermékeként is termelődhetnek. Ezt egy
dsRNS-specifikus nukleáz (Dicer) ismeri fel és hasítja ssRNS-ekké, így
mindkét polaritású, a géncsendesítésre jellemző 21-23 nukleotid hosszúságú kis
RNS molekulák jelen vannak a sejtben, melyek a további RNS degradáció
irányítói (Bass, 2000, Dalmay és mtsai, 2000a, b). Ezek valószínűleg különböző
RNáz-szerű proteinekhez kapcsolódnak, szekvencia-specifikus nukleázokká
alakítva őket, melyek képessé válnak a célszekvenciák felismerésére
(Carrington és mtsai, 2001). A DNS-metiláció szabályozásával közvetve az
aberráns RNS-ek képződésében is szerepet játszanak. Kezdeti lépés lehet
aberráns RNS-ek kialakulása is, mely többféleképpen történhet, pl. metilációval
vagy az ektopikus párosodás (ectopic pairing) jelensége által (Vaucheret és
mtsai, 1998). Ez esetben a transzgének és a homológ endogének közötti
bázispárosodás interferál a transzkripcióval, így nyújtva lehetőséget aberráns
RNS-ek létrejöttére (Baulcombe és English, 1996). Mivel ezek az aberráns

35
RNS-ek bizonyos szintű homológiát mutatnak az eredeti transzkriptumokkal,
szintén templátként szolgálhatnak az RNS függő polimerázoknak, így számos
rövid komplementer szekvenciát képeznek, melyek a funkcionális
transzkriptumokhoz hibridizálhatnak és az így kialakuló kétszálú RNS-eket
specifikus RNázok lebonthatják.
Palauqui és mtsai (1997) oltási kísérleteket végeztek koszupressziót mutató
alanyt és koszupresszió nélküli oltványt használva, mely homológ transzgént
hordozott. A transz- és endogének lecsendesítése minden esetben
bekövetkezett. A jelenséget szisztemikus, szerzett géncsendesítésnek nevezték
el. Megfigyeltek azonban géncsendesítést olyan estekben is, amikor promóter
nélküli transzgénkonstrukcióval (petunia chalcon szintáz génnel)
transzformáltak növényeket. Ez arra utal, hogy maga a szekvenciahomológiát
mutató DNS szakasz is képes indukálni a folyamatot akár szensz akár
antiszensz orientációban (Sijen és Kooter, 2000).
A poszttranszkripcionális géncsendesítés terjedése a vírusok, viroidok
terjedéséhez hasonlóan, a plazmodezmákon és a phloem-en keresztül történik.
A terjedését egy nem-metabolikus, gén-specifikus szignál biztosítja, ami akár
RNS is lehet (Waterhouse és mtsai, 2001b). Mivel a mobilis szignál terjedése a
vírusterjedéssel paralel, így a fertőzés bekövetkezése a szignál és vírus
növényen belüli terjedési idejétől függ. (Vaucheret és Vance, 2001). Ha a vírus
gyorsabb, kialakul a szisztemikus fertőzés, ha a szignálterjedés gyorsabb, akkor
a vírus olyan sejteket talál, ahol a géncsendesítési apparátus már készen áll, így
elmarad a fertőzés. Egyes feltételezések szerint a 21-23 nt RNS-ek nemcsak a
Dicer-komplex degradációjának eredményei, hanem azok a jelek is, amelyek
bejutnak a sejtmagba és a DNS metilációt irányítják, valamint sejtről-sejtre
terjedve az RNS degradációt és a DNS metilációt szabályozzák (Waterhouse és
mtsai, 2001a). Mások szerint az aberráns RNS-ek is részt vehetnek a

36
terjedésben, és a szignál lehet a metilált target gén egy módosított alakja vagy
maga a dsRNS. Oltási kísérletekkel azonban kimutatták (Mallory, 2001), hogy
valószínűleg nem a dsRNS-ek a szignálmolekulák.
A géncsendesítő mechanizmust elsőnek transzgénikus növényekben
figyelték meg, ahol a transzgén és az azzal homológ endogén inaktiválódott.
Mivel a vírusok mind kiváltói, mind célpontjai lehetnek a géncsendesítésnek,
feltételezhető, hogy a poszttranszkripcionális géncsendesítés egy természetes
növényi antivirális reakció. Ez a jelenség egy olyan hatékony mechanizmus,
mely idegen RNS molekulák ellen irányul, amelyek lehetnek ssRNS-ek és
dsRNS-ek és ha egyszer aktiválódott, bármely más, a citoplazmában előforduló
szekvenciahomológiát mutató RNS-ekkel szemben működésbe lép. A tobamo-,
potex- és geminivírusok aktivátorai és célpontjai a géncsendesítésnek megfelelő
nukleinsavhomológia esetén. A caulimo- és nepovírusok a
poszttranszkripcionális géncsendesítéshez hasonló reakciót indukálnak, mint
rezisztenciamechanizmust, akkor is, ha nincs szekvenciahomológia a vírus és a
nukleáris gének között (Ratcliff és mtsai, 1997, Covey és mtsai, 1997). Ezáltal a
szisztemikusan fertőzött levelek tünetmentesekké válnak, lecsökken a
vírusszám és kialakul egy RNS szekvenciaspecifikus rezisztencia a fertőző
vírussal szemben.
A vírusok is kifejlesztettek valamiféle védekező mechanizmust a
poszttranszkripcionális géncsendesítésének elkerülésére vagy gyengítésére.
Megfigyelték pozitív szálú RNS és DNS vírusoknál is, hogy vírusfertőzéskor
korábban lecsendesített gének is reaktiválódhatnak. Néhány vírus olyan
fehérjéket kódol, amelyek képesek a géncsendesítést visszaszorítani, azáltal,
hogy a fertőzéskor képződő szignál létrejöttére vagy mozgására hatnak. Az
eddig felismert szupresszorok a következők: a potyvírusok Hc-Pro fehérjéje, a
potexvírusok p25 fehérjéje, a cucumovírusok 2b-, a sobemovírusok P1-, a

37
tombusvírusok p19- és a geminivírusok Ac2 fehérjéje (Brigneti és mtsai, 1998,
Li és mtsai, 1999, Voinnet és mtsai, 1999, Marathe és mtsai, 2000).
Megemlítendő, hogy ezek a szupresszorok azonban eltérőek evolúciós
eredetükben, szekvenciáikban, struktúrájukban és funkcióikban.
Brigneti és mtsai (1998) GFP-t expresszáló N. benthamiana növényeket
infiltráltak azonos GFP konstrukciót hordozó Agrobacterium tumefaciens-szel,
majd 10 nappal később teljes géncsendesítést tapasztaltak. A koszupresszió
megjelenését követően a növényeket fertőzték PVY-nal, CMV-vel és PVX-szel.
A PVY és CMV esetében a GFP poszttranszkripcionális gén csendesítésének
gátlását tapasztalták a vírusok okozta tünetek mellett, míg a PVX nem hatott a
jelenségre. A mechanizmus vizsgálata érdekében a PVY és a CMV egyes
fehérjéit expresszáltatták PVX vektorban, és kimutatták, hogy a PVY HC-Pro
és a CMV 2b proteinek részt vesznek a poszttranszkripcionális
géncsendesítésének gátlásában. A PVY és a CMV vírusok fehérjéi a növény
eltérő részein szupresszálják a géncsendesítést, így ezek valószínűleg a
folyamat különböző szakaszait blokkolják. Feltételezték, hogy a jelenséget
inkább a proteinek váltják ki, mint az RNS-ek, ugyanis a PVX vektorba épített
módosított, nem transzlálódó szekvenciák nem idéztek elő szupressziót. A HC-
Pro valószínűleg a poszttranszkripcionális gén csendesítés fenntartását gátolja,
míg a 2b protein a szignál bejutását, azaz a jelenség kialakulását akadályozza
meg.
Goodwin és mtsai (1996) TEV CP konstrukcióval transzformált
növényeken végzett vizsgálatok alapján megállapították az RNS által kiváltott
rezisztencia és a transzgén beépülési kópiaszáma közötti összefüggéseket. Azt
tapasztalták, hogy egy vagy két kópiában történő beépülés „recovery”
fenotípust eredményez, három vagy több kópia pedig magas szintű rezisztenciát
vált ki, melyben a poszttranszkripcionális RNS degradáló mechanizmus a vírus

38
eredetű RNS célszekvencia specifikus hasításával lép működésbe, intermedier
termékeket képezve. Azok a növényi vonalak pedig, melyek nyolcnál több
transzgén kópiával rendelkeznek, fogékonyak voltak TEV fertőzéssel szemben.
Ezekben az esetekben a transzgén transzkripcióját nem tudták kimutatni, ami
egy DNS-szintű géncsendesítési mechanizmus jelenlétére és működésére utal.
Mäki-Valkama és mtsai (2000) PVYO törzs P1-génnel transzgénikus Pito
burgonyanövényeket fertőztek PVYN törzzsel, és nem tapasztaltak védettséget,
csupán PVYO fertőzés esetében. A jelenség érdekessége az, hogy a két törzs P1
génjei 96%-nál nagyobb homológiával rendelkeztek és kimutatható struktúrális
eltérés sem volt közöttük. Az eredmények alapján feltételezték, hogy a közel
azonos transzgén- és célszekvenciák ellenére nemcsak a homológiának, hanem
más egyéb faktoroknak is szerepe lehet a géncsendesítés megnyilvánulásában.
2.3.2.5. Kockázati tényezők a köpenyfehérje által mediált keresztvédettség
technológiájának felhasználása esetén
A legtöbb növényi vírus RNS a köpenyfehérjébe csomagolódik be, ez
azonban nemcsak védi azt, hanem részt vesz különféle interakciókban a
gazdanövény és a vírus replikációjának egyes szakaszainál. A CP funkciói
függnek az adott vírustól, ami lehet pl. replikációs szerep (AMV), növényen
belüli mozgás (TMV), felismerési pont a gazdanövény rezisztencia-
mechanizmusaiban (TMV) és részt vehet a vektorokkal történő vírusterjedésben
(Tepfer, 1993).
A CP általi rezisztencia többnyire csak a donorvírussal szemben hatékony,
olykor a közeli rokon vírusok esetében is, de legritkábban a távolabbi rokon
vagy nem-rokon vírusokkal szemben. Ezért egy transzgénikus növényt

39
legvalószínűbben egy távoli rokon vagy nem-rokon vírus fertőzhet meg, így
számolnunk kell a transzgén és a fertőző vírus közötti esetlegesen előforduló
interakciók kialakulásával. A transzgénikus növényekben expresszálódó
vírusgének három különböző jelenség következtében hordoznak magukban
kockázati tényezőt: hetero/transzkapszidáció, rekombináció, szinergista hatás.
2.3.2.5.1. Hetero- és transzkapszidáció
A köpenyfehérje gént expresszáló növényekben egy esetleges rokon
vírustörzs felülfertőződése során kialakulhat a hetero- vagy transzkapszidáció
jelensége, ami annyit jelent, hogy a felülfertőző vírus RNS-ét a transzgénből
származó CP csomagolja be részben vagy egészben, kialakítva a viriont. A
hetero- vagy transzkapszidáció gyakorisága hasonlóan a rekombinációhoz a
kevert fertőzések miatt jól ismert jelenség a növénykórtanban. Az egyik
kockázati tényezőt az jelentheti, ha olyan CP képződik a növényben, amely a
vírus vektorokkal való terjedését segíti és így egy eredetileg vektorral nem
terjedő vírus heteroenkapszidációja lehetővé teszi ennek a vírusnak pl.
levéltetvekkel történő újabb növényre terjedését. A PPV CP-t expresszáló
transzgénikus növényekben ZYMV ’NAT’ törzs levéltetvekkel történő átvitelét
tapasztalták Jaquet és mtsai (1998a), miközben ez az izolátum a saját
köpenyfehérjéje által levéltetvekkel nem képes terjedni. Mivel a vírus
genomjában változás nem történt, az új növényen már az eredeti állapot állt
helyre, azaz ott már nem alakult ki a transzkapszidáció. Ugyanezt a jelenséget
korábban is megfigyelték Atreya és mtsai (1991), Bourdin és Lecoq (1991) ill.
Candelier-Harvey és Hull (1993).

40
Jacquet és mtsai, (1998b) pl. DAG aminosav triplet nélküli CP
transzgénkonstrukció használatával mutatták ki ennek a kockázati tényezőnek
az egyik elkerülési lehetőségét. A fehérje alegységek közül némelyik ugyan
részt vesz a heteroenkapszidációban, azonban a DAG triplet hiányában a
géntermék a levéltetűvel való terjedést nem teszi lehetővé.
Csonkított CP génkonstrukciókkal is sikerült keresztvédettséget kialakítani,
amely konstrukció terméke nem képes a vírus RNS-t becsomagolni, tehát a
vírus nem tud terjedni, miközben a keresztvédettség jelensége megfigyelhető
(Lecoq és mtsai, 1993).
Varrelmann és Maiss (2000) mutáns PPV CP géneket építettek
fertőzőképes, teljes hosszúságú PPV klónba, melyben megváltoztatták a virion
összeszerelődéséhez szükséges (R3015Q3016 és D3059) motívumokat. Az így
létrehozott konstrukciókkal fertőztek transzgénikus, vad típusú PPV CP-t
expresszáló- és nem transzgénikus N. benthamiana növényeket. A nem
transzgénikus növényekben nem tapasztaltak vírusterjedést és a virionok sem
alakultak ki, míg a CP-t expresszáló növényekben megfigyelték a vírus
szisztemikus mozgását és a víruspartikulumok összeszerelődését. A vad típusú
vírus CP-t tartalmazó fertőzőképes klón esetében a nem transzgénikus
növényekben azonban megfigyelték a vírus mozgását, ezzel bebizonyították,
hogy a mutáns klónok esetében a szisztemikus fertőzés hiányát egyedül a vírus
CP összeszerelődési motívumainak megváltoztatása okozta. Ezeket az
eredményeket felhasználva létrehoztak transzgénikus N. benthamiana
növényeket, melyek a mutáns CP-ket expresszálták. A korábbi megfigyeléseket
igazolva a transzgénikus növények fertőzését követően a heteroenkapszidáció
kialakulásának jelentős csökkenését állapították meg a transzgén eredetű CP-k
és a fertőző potyvírusok köpenyfehérjéi között.

41
A különböző kísérletekben felvázolt transzgén konstrukciók (melyek
különféle köpenyfehérje modifikációkat kódolnak) megfelelő és körültekintő
alkalmazásával a hetero- és transzkapszidáció veszélye bizonyíthatóan
minimálisra csökkenthető.
2.3.2.5.2. Homológ és heterológ rekombináció
Vírus eredetű köpenyfehérje gént expresszáló növény esetleges
felülfertőződése rokon vírussal nem különbözik a természetben is meglévő
fogékony növényben előforduló két vagy több vírussal történő szimultán
fertőződéstől. Ezt a jelenséget minden egyes vírus esetében megtaláljuk, hiszen
szinte kivétel nélkül több vírustörzs van jelen egy fertőzött növényben. Irodalmi
adatok szerint egy természetes vírusfertőzés esetén például a polio- és
influenzavírusoknál a rekombináció elérheti a 35%-os gyakoriságot, a növényi
vírusoknál azonban ez a szám nagyságrendekkel kisebb. Azonosításra kerültek
a CP gént kódoló RNS 3’ végén elhelyezkedő rekombinációs „forró pontok”,
melyek eltávolítása mellett is sikerrel lehet olyan konstrukciókat előállítani,
amelyekkel a keresztvédettség indukálható. A CP gént expresszáló
növényekben, ha azok ellenállóak, elvileg vírussal nem fertőzhetők felül, így a
rekombináció sem alakulhat ki. Falk és mtsai (1994) szerint a korábbiaktól
eltérő, új károkat okozó vírusok nem a transzgénekkel történő
rekombinációkból eredhetnek, hanem a már meglévő vírusgenomok közötti
variációkból. A vírusevolúció szerves részének kell tekintenünk a
rekombinációt, melynek gyakorisága a természetben igen alacsony értéket
mutat, vagyis nehezen mutatható ki, mivel a replikáció alatt leggyakrabban
homológ szakaszok között jön létre (Bruyere és mtsai, 2000). Kísérleti adatok

42
szerint a transzgénikus növényekben előforduló rekombináció gyakorisága
messze elmarad a természetben előforduló vírusfertőzések esetében
kialakulóktól (Vö. Tepfer és Balázs 1997).
Chiang és mtsai (2001) különböző összetételű rekombináns CP gént
hordozó PRSV-vel és PRSV-HA CP génnel transzgénikus papaya növényeken
végeztek kísérleteket a vírusok virulenciájának vizsgálatára. Megfigyelték,
hogy a vírus fertőzőképessége nagyobb mértékben függ a transzgén CP és a
vírus kiméra CP-je közötti szekvenciaazonosság helyzetétől, mint a fokától. A
rekombinánsok virulenciájának eltéréseire magyarázatként szolgálhat a
géncsendesítés jelensége. Megállapították, hogy a PTGS elsősorban a transzgén
3’ vég régióját célozta meg, ezért ha a vírus CP génje és a transzgén 3’ régiója
homológ, akkor a fertőzés enyhébb, vagy el is maradhat.
2.3.2.5.3. Szinergizmus
A növénykórtanban jól ismert az a jelenség, amikor két vagy több vírus
ugyanazon növényegyeden történő együttes fertőzésekor kialakuló tünetek
súlyosabbak, mint a vírusok külön-külön okozott tünetei. Jól példázza ezt a
burgonya X vírus és Y vírus együttes fertőzése esetén a növény igen súlyos
kórképe, amely akár teljes pusztuláshoz is vezethet. A jelenség érdekessége az,
hogy a burgonya X vírus egyedüli fertőzés esetén szinte tünetmentes a
burgonyanövényen. Nem tudjuk még, hogy egyes vírusgének megnyilvánítása a
kultúrnövényekben egy felülfertőző vírussal együttesen kialakíthat-e ilyen
szinergista hatást, azaz a betegség kórképének megváltozását, vagy a növény
esetleges pusztulását. Ennek a rizikótényezőnek a kísérletes tanulmányozása
mindenképpen napjaink és a jövő feladata.

43
3. ANYAG ÉS MÓDSZER
3.1. ANYAGOK
3.1.1. Növényi anyag
A transzformációs kísérletben felhasznált növények a Magyarországon
régebben elterjedt, ma már érzékenységük miatt a köztermesztésből kiszorult,
steril körülmények között, táptalajon nevelt Solanum tuberosum L. cv Mindenes
(M) és Somogyi Kifli (SK) magyar burgonyafajták, melyek a Veszprémi
Egyetem Georgikon Mezőgazdaságtudományi Kar Burgonyakutatási
Osztályának génbankjából, Keszthelyről származnak.
A Nicotiana tabacum Virgin D (VD), Stamm C2 (SC) és Hevesi 11 (H11)
dohányfajták az AGROTAB Nemesítő és Vetőmagtermeltető Kft-ből
(Debrecen- Pallag) származnak. A Virgin D német fajta, a Stamm C2 német
nemesítési vonal, míg a Hevesi 11 magyar fajta, melytől az állami elismerést
visszavonták a vírusérzékenysége miatt. Mindhárom fajta rendelkezik bizonyos
szintű genetikai vírus-ellenállósággal a PVY-nal szemben, azonban ez egy
erősebb fertőzöttségű évben nem nyújt megfelelő védelmet.
A burgonyanövények fenntartása steril hajtáscsúcs-tenyészetek formájában,
RB táptalajon (Ishida és mtsai, 1989), míg a dohánynövények fenntartása
hasonlóképpen, de RM táptalajon történt, fényszobában, kontrollált
körülmények között (23 oC, 16 óra megvilágítás).

44
3.1.2. Vírusizolátum
A kísérletben használt PVY-H a nekrotikus törzsek közé tartozó magyar
izolátum, Dr. Beczner Lászlótól származik (Növényvédelmi Kutatóintézet,
Budapest), melynek jellegzetessége, hogy a burgonyagumón és a bogyón
nekrotikus gyűrűsfoltosságot okoz, illetve rezisztencia áttörő képességgel is
rendelkezik. Az izolátum patológiailag (Beczner és mtsai, 1984) és
molekulárisan is jellemzett (Thole és mtsai, 1993).
A PVY-H vírustörzs folyamatos fenntartásához és felszaporításához
Nicotiana tabacum cv Xanthi-nc dohányfajtát használtunk.
3.1.3. Génkonstrukció
A növények transzformálásához az MBK virológia csoportjában korábban
előállított génkonstrukciót tartalmazó pGAYHCP plazmidot használtuk (Kollár
és mtsai, 1993). A pGA482 növényi expressziós vektor alapú
génkonstrukcióban az AUG transzlációs start kodonnal ellátott PVY-H
köpenyfehérje gén a karfiolmozaik vírus (CaMV) 35 S promóter és a NOS
terminációs szabályozórégiók között helyezkedik el. A plazmid tartalmaz
tetracyclin és kanamycin rezisztenciagéneket is (6. ábra).

45
6. ábra
A transzformációkhoz használt génkonstrukció sematikus ábrája
3.1.4. Baktérium törzs
A növényi transzformációhoz a Rhizobiaceae családba tartozó Gram-
negatív Agrobacterium tumefaciens talajbaktérium C58C1 (Rifr) törzsét
használtuk (Zambryski és mtsai, 1983). A pGAYHCP plazmidot tartalmazó
Agrobacterium törzset antibiotikumokkal kiegészített YEP táptalajon tartottuk
fenn. Átoltáskor 2-3 napig 28 oC-on, majd 4 oC-on tároltuk. A növényi
transzformációt megelőzően pedig folyékony CPY tápoldatban szaporítottuk
fel. 12-24 óráig, 30 oC-on rázatva 180-200 fordulat/perc.

46
3.1.5. Táptalaljok, tápoldatok
CCC táptalaj (Ishida és mtsai, 1989): /1000ml/:50ml MSI (20x makro) /Murashige and Skoog, 1962/, 5ml MSII (200x mikro),5ml MSIII, 5mg BA, 500mg CCC /klór-kolin-klorid/, 2mg glicin, 0.5mgnikotinsav, 0.5mg piridoxin, 0.4mg thiamin, 100mg mezoinozit, 80g szacharóz,6.5g agar, pH: 5.8.
CPY tápoldat /1000ml/: 1g Bacto yeast extract, 5g pepton vagy kazeinhidrolizátum, 5g szacharóz.
G80 táptalaj /1000ml/:50ml MSI, 5ml MSII, 5ml MSIII, 2.5mg thiamin, 12.5mg piridoxin, 12.5mgnikotinsav, 10mg AgNO3, 50mg glicin, 0.5mg folsav, 0.05mg biotin, 100mgmezoinozit, 30g szacharóz, 6.5g agar, pH: 5.7, 500mg cefotaxim, 80mgkanamycin.
MSS táptalaj /1000ml/:50 ml MSI, 5 ml MSII, 5ml MSIII, 100mg mezoinozit, 0.4g thiamin, 0.5mgnikotinsav, 0.5mg piridoxin, 1mg BAP, 1mg NAA, 30g szacharóz, 6.5g agar,pH:5.8
MSR táptalaj /1000ml/:50ml MSI, 5ml MSII, 5ml MSIII, 100mg mezoinozit, 0.4g thiamin, 0.5mgnikotinsav, 0.5mg piridoxin, 2mg glicin, 1mg IAA, 20g szacharóz, 6.5g agar,pH:5.8
MSI /1000ml/:33g NH4NO3, 38g KNO3, 8.8g CaCl2.H2O, 7.4g MgSO4, 3.4g KH2PO4.
MSII /500ml/:0.083g KJ, 0.062g H3BO4, 0.223g MnSO4.4H2O, 0.025g NaMo4.2H2O, 0.086gZuSO4.7H2O, 0.0025g CuSO4.5H2O, 0.0025g Cl2.6H2O.
MSIII /100ml/:0.55g FeSO4.7H2O, 0.74g Na2EDTA.
RB táptalaj (Ishida és mtsai, 1989): /1000ml/ 50ml MSI, 5ml MSII, 5ml MSIII, 0.4mg thiamin, 100mg mezoinozit, 30gszacharóz, 6.5g agar, H2O, pH: 5.7

47
RB1 táptalaj (Ishida és mtsai, 1989): /1000ml/ : 50ml MSI, 5ml MSII, 5ml MSIII, 0.4mg thiamin, 100mg mezoinozit, 20gszacharóz, 6.5g agar, pH: 5.7
RB2 táptalaj (Ishida és mtsai, 1989): /1000ml/: 50ml MSI, 5ml MSII, 5ml MSIII, 2.5mg thiamin, 12.5mg piridoxin, 12.5mgnikotinsav, 50mg glicin, 0.5mg folsav, 0.05mg biotin, 100mg mezoinozit,0.03mg NAA (naftil-ecetsav), 1mg BA, 0.5mg zeatin, 10mg AgNO3, 30gszacharóz, 6.5g agar, pH: 5.8, 50mg kanamycin, 500mg cefotaxim.
RM táptalaj /1000ml/:50ml MSI, 5ml MSII, 5ml MSIII, 20g szacharóz, 6.5g agar, pH:5.8.
YEP táptalaj /1000ml/: 10g Yeast Extract (élesztőkivonat), 10g pepton, 5g NaCl, 15 g agar, pH: 7.0,25mg kanamycin, 100mg rifampicin, 5mg tetracyclin.
3.1.6. Oldatok, pufferek
Prehibridizáló és hibridizáló oldat (Sambrook és mtsai,1989):5x SSPE, 5x Denhard's oldat, 0.5%-os SDS, 1mg Yeast RNA, 50% H2O.
Extrakciós puffer /60ml/:3ml 1M Tris-HCl, 1.2ml 5M NaCl, 120µl 0.5M EDTA, 3ml 10% SDS, 2-ME(merkapto-etanol) 0.3%, pH:7.6.
Hibridizációs mosóoldat:100ml 20xSSC, 5ml 10%-os SDS, 895ml H2O.
PCR reakció:templát DNS (10µl-ben), 3 µl MgCl2 (25mM) USB, 5 µl Buffer (10x), 2 µldNTPs (5mM), 1-1 µl oligonukleotid (0.15 µg/ µl), 2.5 unit Taq polimeráz, 27.5µl H2O.
10x TBE puffer /1000ml/:121.1g Tris, 51.35g bórsav, 3.72g EDTA-Na, pH: 8.3.

48
20xSSC oldat /1000ml/:175,3g NaCl, 88,2g Na-citrát, pH: 7.0.
3.2. MÓDSZEREK
3.2.1. Módszerek transzgénikus növények előállításához
3.2.1.1. Növény transzformáció
A növények felszaporításához és transzformációjához steril tenyészetekből
indultunk ki.
A PVY-H CP gént tartalmazó PGAYHCP plazmidot Agrobacterium
tumefaciens segítségével építettük be burgonya- és dohánynövényekbe. A
transzformációhoz 28 oC-on, egy éjszakán át növesztett baktériumkultúrát, kb
0.5-1 cm-es levélkorongokat és szárszegmenseket, valamint burgonya esetében
0.5-1 cm-es mikrogumók 2-3 mm-es korongjait használtuk (Ishida és mtsai,
1989). A levéldarabkák esetében a sebzési felületet steril szikével történő
bemetszésekkel növeltük. A növényi részeket petricsészébe helyeztük
antibiotikummentes RM (dohány), illetve RB1 (burgonya) táptalajra, majd ezek
felületére pipettával cseppentve vittünk fel 5-20 µl folyékony CPY tápoldatban
felszaporított Agrobacterium tenyészetet. A 2 napon át fényszobában,
kontrollált körülmények között inkubált mikrogumó korongokat illetve a levél-
és szárszegmenseket a baktérium elölése céljából 0.5-3 órán át 500mg/l
cefotaxim oldatban mostuk, majd megszárítottuk.

49
3.2.1.2. Hajtásindukálás
A lemosott, majd megszárított mikorgumó korongokat, szár- és
levélszegmenseket kanamycint (300mg/l) és cefotaximot (500mg/l) tartalmazó
RB2 táptalajra helyeztük burgonya esetében (Ishida és mtsai, 1989), míg
dohánynövényeknél MSS táptalajra és fényszobában 23 oC-on, 16 órás
megvilágítás mellett neveltük. A tenyészeteket 2 hetente új RB2 illetve MSS
táptalajra helyeztük át. 4-5 hét elteltével jelentek meg a hajtások a mikrogumó
korongok illetve a levél- és szárszegmensek szélein.
3.2.1.3. Gyökereztetés
Két- három hét elteltével az 1-2 cm-es méretet elért burgonyahajtásokat
steril körülmények között leválasztottuk a mikrogumó-, levél- vagy
szárdarabkákról, majd G80, 80 mg/l kanamycint tartalmazó ún. gyökereztető
táptalajba helyeztük 100ml-es Erlenmeyer lombikba, illetve "Magenta" és
VEGBOX® műanyag dobozokba. A mikrogumó-, levél- és szárszegmenseket
friss RB2 táptalajra helyeztük vissza, újabb hajtásnövekedést indukálva.
Dohánynövényeknél ugyanezen célból az MSR, 150 mg/l kanamycint
tartalmazó táptalajt használtunk.
3.2.1.4. Kiültetés üvegházba
Megfelelő méretű gyökérzet kifejlődésekor a maradék táptalajt
leválasztottuk, majd lemostuk a növényekről, és a gyökeres hajtásokat steril
földbe ültettük, illetve üvegházi körülmények között neveltük tovább.

50
Folyamatos fenntartásukat egyrészt fényszobában, másrészt üvegházban
önmegtermékenyítéssel és magfogással a dohányfajtáknál illetve a
burgonyafajtáknál az előző évben megtermelt gumóik folyamatos elültetésével
biztosítottuk.
3.2.2. Transzgénikus növények vizsgálata
3.2.2.1. A köpenyfehérje gén integrációjának vizsgálata
A transzgén beépülését a növényi genomba PCR-rel (polymerase chain
reaction) vizsgáltuk. A növényi DNS kivonása Rubino és mtsai (1992) által leírt
módszer szerint történt. A kb. 200mg-os levéldarabkákat 400 µl extrakciós
pufferrel dörzscsészében eldörzsöltük. 2 µg kivont növényi DNS-t és két, a
PVY CP génre és a 3 NTR-re tervezett oligonukleotid primert használtunk,
melyek a 8776-8800 illetve a 9376-9394 nukleotid szakaszokkal homológok:
YCP 2 oligonukl. pr.: 5’-CACTTGCTCGAGTATGCTCCACAGC- 3’,
YCP 4 oligonukl. pr.: 5’-CGTCCGGAGAGACACTACA –3’.
A primerek egy 618 bp-nyi DNS szakasz kiemelését tették lehetővé. Az 5
perces 94 oC-os denaturáló lépést 35 ciklus követte (15 sec 94 oC, 30 sec 60 oC,
90 sec 72 oC). Az utolsó polimerizáló lépés 10 percig tartott. Mindezt Perkin
Elmer Cetus / DNA Thermo File készülékkel végeztük.
A PCR termékeket gél elektroforézissel és Southern hibridizációval
(Sambrook és mtsai, 1989) ellenőriztük: a PCR termékeket 1:1 arányban
kloroformmal extraháltuk, majd 10 µl-t mintánként 1%-os agarózt tartalmazó
TBE gélben megfuttattunk 120 V feszültség mellett. A DNS-t 20xSSC oldat
által Hybond-N membránra (Amersham) blottoltuk, majd a DNS megkötődése
érdekében a membránt 3 percre UV fény alá helyeztük.

51
A prehibridizálás és a hibridizálás 65 oC-on, a gyártó (Amersham)
utasításai szerint történt. A próba jelölése T7Quick Prime Kit alapján, random
priming úton történt a gyártó (Pharmacia Biotech) előírásai szerint. A jelöléshez
17 µl 25-50ng-ot tartalmazó pGAPVYHCP plazmidot és α-32P dCTP izotópot
használtunk, majd másnap hibridizációs mosóoldatban történő többszöri mosás
után a membránról autoradiogramot készítettünk.
3.2.2.2. Transzgén expresszió vizsgálata
Az RNS átíródását Northern analízissel vizsgáltuk. A növényi
totálnukleinsavat White és Kaper (1989) által leírt módszer szerint izoláltuk,
formaldehidet tartalmazó gélen elválasztottuk, majd Hybond-N membránra
(Amersham) blottoltuk. A mintákat ezután α-32P dCTP izotóppal jelölt próbával
hibridizáltattuk 42 oC-on, az Amersham által megadott leírás szerint.
A géntermék megjelenésének kimutatására Western analízist (Sambrook és
mtsai, 1989) végeztünk. A levélből kivont összes proteint 12 %-os
poliakrilamid gélen elválasztottuk, majd Hybond-C extra membránra
(Amersham) blottoltuk. A megkötött fehérjék kimutatásához a kereskedelmi
forgalomban kapható alkalikus foszfatázzal konjugált anti- PVY CP antitestet
(Boehringer Mannheim GmbH) és kromogenikus szubsztrátjait (5-bromo-4-
chloro-3-indolyl-phosphatepara-toluidine-t és para-nitro-blue-tetrazolium-
chloride) használtuk.

52
3.2.2.3. Rezisztenciavizsgálatok
3.2.2.3.1. A mesterséges fertőzés és körülményei
A transzgénikus dohánynövényeket tisztított PVY-H -val fertőztük az
ellenállóképesség vizsgálatának céljából. 0.1M-os Na-foszfátpufferrel (pH:7.2)
hígított 0.5 µg/ml, 5 µg/ml és 20µg/ml víruskoncentrációt használtunk a
dohányvonalak fertőzéséhez (1.táblázat). A Virgin D dohányfajta esetében 6
vonalat, az Stamm C2 és a Hevesi 11 fajtáknál 12-12 vonalat fertőztünk.
Vonalanként 3-3 dohánynövényt használtunk egy-egy víruskoncentrációhoz. Az
alsó két levelet fertőztük mechanikailag, carborundum por és üvegspatula
segítségével. A mintavétel a fertőzést követő 5., 10., 15., 20. és 25. napon
történt a növények felsőbb leveleiből. A minták kiértékeléséhez dot-blot
analízist végeztünk. A fertőzést követően a növényekből kivont RNS-t Hybond-
N membránra (Amersham) csepegtettük kétféle koncentrációban. A
membránról α-32P dCTP izotóppal jelölt CP specifikus próbával történő
hibridizációt követően autoradiogramot készítettünk.

53
1. táblázat
Az üvegházi rezisztenciavizsgálatok paramétereinek összefoglaló táblázata.
Megfertőzött
dohányvonalak
Fertőzéshez használt
víruskoncentráció
(PVY-H)
Mintavétel ideje
(dpi)
Virgin D: 6 vonal
Stamm C2: 12 vonal
Hevesi 11: 12 vonal
0.5 µg/ml
5 µg/ml
20 µg/ml
5,
10,
15,
20,
25.
3.2.2.3.2. Szabadföldi kísérletek
22 Mindenes és 4 Somogyi Kifli transzgénikus burgonyavonalat vontunk
be a szántóföldi vizsgálatokba. Ezek egy részével 1999-ben kezdődtek a
kísérletek, míg egy másik csoporttal 2000-ben.

54
1999-ben vizsgálatba vont vonalak: Mindenes 2, 3, 4, 5, 6, 7,
10, 11, 12, 21, 24.
Somogyi Kifli 1, 2.
2000-ben vizsgálatba vont vonalak: Mindenes 1, 8, 9, 14, 15,
16, 17, 18, 20, 27, 33.
Somogyi Kifli 3, 4.
A szántóföldi vizsgálatok első lépése a vonalak in vitro vírusmentesítése
volt, ezt követte a felszaporításuk. Az 1999-es évben vizsgálatokba vont
vonalaknál megkezdett kísérleti módszerek a következők voltak:
- mesterséges mechanikai fertőzés PVYNTN D-10 izolátummal,
ELISA teszt (2000),
- szabadföldi infektor növényes vizsgálat (2000),
- kertészeti oltás (2001),
- morfológiai-, gazdasági- és felhasználhatósági értékvizsgálatok (2001).
A 2000-ben szántóföldi vizsgálatokba vont vonalaknál is vírusmentesítés
volt az első lépés. Ezeknél a vonalaknál az eddigi vizsgálati módszerek a
következők voltak:
- mesterséges mechanikai fertőzés PVYNTN D-10 és PVYO/C Ka-49
izolátumokkal, ELISA vizsgálat (2001),
- Solanum demissum A6 bioteszt (2001).
Ezen vonalak felszaporítása és további, több ismétléses vizsgálata jelenleg is
folyik.

55
3.2.2.3.2.1. Kisparcellás, infektor növényes, szabadföldi kísérlet
A transzgénikus burgonyavonalak szántóföldi vizsgálatát Dr. Horváth
Sándorral és Dr. Wolf Istvánnal együttműködve, a Veszprémi Egyetem
Georgikon Mezőgazdaságtudományi Kar Burgonyakutatási Központtal
(Keszthely) végeztük. A transzgénikus vonalak szabadföldre, kisparcellákba
kerültek kiültetésre, kukorica szegélyvetéssel, mely körbevette az egész
kísérleti területet. Soronként 20 gumó elültetése történt, vonalanként egy illetve
két sorban. Egy sor 5 m hosszú volt. Negatív kontrollként vírusmentes, nem
transzgénikus Somogyi Kifli és Mindenes sorok, illetve mechanikailag
PVYNTN-nel inokulált Somogyi Kifli sorok, mint infektor növények
helyezkedtek el a transzgénikus sorok között. A növények kisparcellán belüli
elrendezését a 7. ábra szemlélteti.

56
fertőzött Somogyi Kifli
Somogyi Kifli 2- dupla sor
fertőzött Somogyi Kifli
vírusmentes Somogyi Kifli kontroll- dupla sor
fertőzött Somogyi Kifli
Mindenes 3- dupla sor
fertőzött Somogyi Kifli
Mindenes 5
Mindenes 6
fertőzött Somogyi Kifli
Mindenes 10- dupla sor
fertőzött Somogyi Kifli
vírusmentes Mindenes kontroll- dupla sor
fertőzött Somogyi Kifli
Mindenes 11- dupla sor
fertőzött Somogyi Kifli
Mindenes 12
Mindenes 21
fertőzött Somogyi Kifli
Mindenes 21
fertőzött Somogyi Kifli
7. ábra
A transzgénikus burgonyavonalak és kontrollok elrendezése
a kisparcellás szabadföldi kísérletben.

57
3.2.2.3.2.2. ELISA vizsgálat
A növényi vírusok detektálására a módszer egyszerűsége és
megbízhatósága miatt a double antibody sandwich (DAS) ELISA (Enzyme
Linked Immunosorbent Assay) technika alkalmazására került sor (Clark és
Adams, 1977). A módszer előnye, hogy nagy érzékenységű, nanogram
mennyiségű vírusfehérje jelenléte is kimutatható.
A transzgénikus Mindenes és Somogyi Kifli burgonyagumókból kifejlődő
növények 4-leveles állapotában történt a mechanikai fertőzés (karborundum
por - üvegspatula módszerrel). A fertőzést követő 36. napon történt a vizuális
felvételezés, a 37. napon pedig a DAS-ELISA vizsgálatok. A felhasznált
antitestek a Boehringer Mannheim Gmbh illetve a Loewe Biochemia Gmbh
által gyártott poliklonális antitest- és alkalikus foszfatázzal konjugált antitest
alapú kitek voltak.
Vírusizolátum: PVYNTN, D-10, származás: Dunaegyháza burgonya
PVYO/C, Ka-41, származás: Kaba burgonya
(Wolf és Horváth, 2000)
Présnedvhígítás: 1/10, 1/15
Antitest hígítás: 1/1000 (150 µl/vályat)
Lemez: Corning Costar, Cat No: 3590.
Az extinkciós érték mérése EPD-1 fotométerrel, 405 nm-en történt.

58
3.2.2.3.2.3. Solanum demissum A6 teszt
A Solanum demissum A6 a Solanum demissum x S. tuberosum cv Aquilla
fajta származéka, mely biotesztekhez elterjedten használatos. A PVY minden
törzsével szemben nekrotikus lokális léziókkal reagál (in: Takács, 2001). A S.
demissum A6 teszt leválasztott S. demissum A6 levelek fertőzésével történik. A
vizsgálathoz vonalanként 4-4 burgonyanövény és növényenként 2-2 S.
demissum A6 levél fertőzése történt a transzgénikus növények szövetnedvével
azok PVYNTN és PVYO/C fertőzését követő 37. napján. A leválasztott és
inokulált levelek „nedves kamrában”, azaz petricsészében, nedves
szűrőpapíron, szobahőmérsékleten inkubálódtak. A vírusfertőzés tüneteinek
vizsgálata a 6-7. napon történt.
3.2.2.3.2.4. Kertészeti oltás kísérletek
A szabadföldi vizsgálatok eredményei alapján kiválasztott Mindenes 11, 12
és 21-es vonalak kerültek kertészeti oltással történő kísérletekbe. A PVYNTN-nel
mesterségesen fertőzött dohány- és paradicsom alanyra történt a transzgénikus
burgonyanövények oltása, illetve transzgénikus burgonya alanyra a fertőzött
paradicsom oltóág. A kísérletekben azonos korú, 8-10 leveles alany és oltóág
vett részt. Dohány alany esetében vonalanként 2-2 növény, paradicsom- és
burgonya alany esetében 3-3 növény vizsgálata történt meg. A
víruskoncentrációt az ELISA vizsgálatok eredményei mutatták az oltást követő
4. héten.
Határérték kiszámítása mutatta, hogy egy növény mikor tekinthető
fertőzöttnek: Az extinkciós értékekből lettek kivonva a vak oldat (csak puffer)
értékek. Az így kapott maximum és minimum érték különbségének a

59
háromszorosához kell hozzáadni a negatív kontrollok (nem fertőzött növények)
átlagértékét, és az így kapott érték a fertőzési határérték, mely alatt a növény
nem tekinthető fertőzöttnek, míg ennél magasabb érték esetében vírusfertőzött.
Az eredmények megerősítése S. demissum A6 bioteszttel történt.
3.2.3. Vírusmentesítés
A transzgénikus burgonyavonalak szántóföldi vizsgálatait megelőző
vírusmentesítés szövettenyésztés, hőterápia és kemoterápia együttes
alkalmazásával történt. A gumóról leválasztott hajtáscsúcs (nem merisztéma)
MS táptalajon inkubálódott, majd miután egész növénnyé fejlődött, vonalanként
10 nodális szegment 30 mg/l virazolt (ribavirin) tartalmazó táptalajra került,
majd 2 hétig 21 oC-on 16/8 órás fény/sötét periódus mellett nevelkedtek. 2 hét
elteltével a szegmentek 4 hétre hőterápiás kamrába kerültek, ahol 4 óra sötét 32oC és 4 óra fény 37 oC váltakozott. 4 hét után minden növény hajtáscsúcsa
visszakerült MS táptalajra, 21 oC-os és 16 órás megvilágítottságú körülmények
közé.
3.2.4. Vírustisztítás
A transzgénikus növények fertőzéséhez a tisztított vírust Leiser és Richter
(1978) által leírt módszer néhány módosításával nyertük. A részletes leírás
Thole (1993) kandidátusi értekezésében található.

60
4. EREDMÉNYEK
4.1. TRANSZGÉNIKUS NÖVÉNYEK ELŐÁLLÍTÁSA
A PVY-H CP gént tartalmazó pGAYHCP plazmidot Agrobacterium
tumefaciens segítségével építettük be burgonya- és dohánynövényekbe. A
transzformációhoz levélkorongokat, szárszegmenseket és burgonya esetében
mikrogumó korongokat is használtunk, melyeket kanamycint (300mg/l)
tartalmazó táptalajon szelektáltunk. A transzformációt követő 4-6 héten jelentek
meg a hajtások (8/A ábra). Kontrollkísérletek elvégzésével igazoltuk, hogy a
szelektív táptalajon csak a transzformált növényi részeken történik
hajtásnövekedés. A transzformánsokat 150mg/l kanamycint tartalmazó
táptalajon gyökereztettük (8/B ábra) fényszobában (16h megvilágítás / 23 °C),
majd üvegházban földbe ültettük és neveltük tovább (8/C ábra).
A Virgin D dohányfajta esetében 38, Stamm C2 esetében 55 és Hevesi 11
esetében 23 transzgénikus vonalat kaptunk. A Mindenes burgonyafajtából 37
vonalat és a Somogyi Kifli fajtából 4 vonalat tudtunk előállítani.
A kapott transzgénikus vonalak mindegyikét vizsgáltuk molekuláris
módszerekkel, a dolgozatban azonban nem mutatjuk be részletesen mind a 116
dohány- és 41 burgonyavonal molekuláris vizsgálatainak eredményét, ezek
összefoglalása a 8. és 9. táblázatban (74. o.) található. Ábráinkon egyes
kiválasztott, reprezentatív vonalak vizsgálatainak pozitív eredményeit követjük
nyomon.

61
8. ábraA: Mikrogumó korongon, steril tenyészetben fejlődő transzgénikus
burgonya hajtásB: G80 táptalajon nevelt gyökeresedő transzgénikus burgonyanövényC: Üvegházi körülmények között nevelt transzgénikus burgonyanövény
A B
C

62
4.2. TRANSZGÉNIKUS NÖVÉNYEK VIZSGÁLATA
4.2.1. Az integrálódott CP gén kimutatása
A transzgén növényi genomba épülését PCR-rel, majd ezt követően
Southern analízissel vizsgáltuk (9. és 10. ábra). Az adott génre specifikus,
radioaktívan jelölt próbával végzett hibridizáció megerősítette a PCR termékek
CP génnel való azonosságát. A további molekuláris vizsgálatokat a pozitív PCR
eredmények ismeretében végeztük.
9. ábra
Southern blot transzgénikus dohánynövényekből32P dCTP izotóppal jelölt CP specifikus próbával hibridizáltatott transzgénikusdohánynövények PCR termékeinek autoradiogramja. 1. sáv: VD1, 2. sáv: VD2,3. sáv: VD3, 4. sáv: VD4, 5.sáv: VD5, 6. sáv: VD6, 7. sáv: nem transzgénikusdohánynövény, mint negatív kontroll, 8. sáv: PVY-nal fertőzött, nemtranszgénikus dohánynövény, mint pozitív kontroll.

63
10. ábra
PCR termékek transzgénikus burgonyanövényekből és ezek Southern blot-ja
Burgonyanövények PCR termékei (felső ábra), illetve ezek 32P dCTP izotóppaljelölt CP specifikus próbával hibridizáltatott autoradiogramja (alsó ábra). 1.sáv: nem trg. Mindenes kontroll, 2. sáv: Mindenes 2, 3. sáv: Mindenes 3, 4. sáv:Mindenes 4, 5.sáv: Mindenes 5, 6. sáv: Mindenes 6, 7. sáv: Mindenes 7, 8. sáv:Mindenes 8, 9. sáv: fertőzött növény, mint pozitív kontroll, 10. sáv: negatívkontroll (víz), 11 sáv: méretmarker.
4.2.2. Transzgén expresszió vizsgálata
A fehérjetermék kimutatásához Western analízist végeztünk.
A dohánynövényeknél a Virgin D fajta 38 vonala esetében az összes
pozitívnak bizonyult, a Stamm C2 fajta 55 vonalából egyértelműen pozitív 26
vonal, gyenge jelet kaptunk 8 vonal esetében. A Hevesi 11 fajta 23 vonalában
pedig 17 volt egyértelműen pozitív. Negatív kontrollként nem transzformált,

64
egészséges növény összfehérje kivonatát választottuk el a gélen, pozitív
kontrollként pedig fertőzött nem transzgénikus növényt használtunk (11. ábra).
11. ábra
Western blot dohánynövényekből
Transzgénikus növények fehérjekivonataiból kimutatható köpenyfehérje. 1 sáv:pozitív kontroll, fertőzött, nem transzgénikus növény, 2. sáv: negatív kontroll,3.sáv: VD2, 4. sáv: VD3, 5. sáv: VD4, 6. sáv: VD5, 7. sáv: VD6, 8.sáv: VD7, 9.sáv: VD8, 10. sáv: VD9.
A burgonyafajtáknál a Mindenes 37 vonala esetében 34 vonal termelte a
köpenyfehérjét, 3 vonal pedig nem (12. ábra). A Somogyi Kifli fajta 4
vonalából 3-ban tapasztaltunk köpenyfehérje termelődést, ebből 1 esetben
alacsony szinten, míg a 4. vonal egyáltalán nem termelte a transzgén terméket.

65
12. ábra
Western blot burgonyanövényekből
Transzgén eredetű köpenyfehérje kimutatása membránhoz kötötten. 1. sáv:pozitív kontroll, fertőzött, nem transzgénikus növény, 2. sáv: negatív kontroll,3. sáv: Mindenes 2, 4. sáv: Mindenes 3, 5. sáv: Mindenes 4, 6. sáv: Mindenes 5,7. sáv: Mindenes 7, 8. sáv: Mindenes 8, 9. sáv: Mindenes 9, 10. sáv: Mindenes10.
4.2.3. Rezisztenciavizsgálat üvegházban, dot-blot analízis
A Virgin D dohányfajta esetében 6 vonalat, az Stamm C2 és a Hevesi 11
fajtáknál 12-12 vonalat fertőztünk PVY-H-val. Mindhárom használt
víruskoncentráció esetén (0,5µg/ml, 5µg/ml, 20µg/ml) 3-3 növényt, azaz a VD
fajtánál összesen 54 növényt, SC és H11 fajtánál 198-198 növényt vizsgáltunk
üvegházi körülmények között.
A Mindenes burgonyafajta esetében 22 vonalat, a Somogyi Kifli fajtánál 3
vonalat fertőztünk 5 µg/ml-es víruskoncentrációval.

66
Az üvegházi rezisztenciavizsgálatokban használt transzgénikus dohány- és
burgonyavonalak:
Virgin D (dohány): VD1, VD2, VD3, VD4, VD5, VD6.
Stamm C2 (dohány): SC35, SC36, SC39, SC40, SC41, SC43,
SC44, SC45, SC46, SC47, SC48, SC49.
Hevesi 11 (dohány): H11-1, H11-2, H11-3, H11-4, H11-6,
H11-7, H11-8, H11-9, H11-10, H11-11,
H11-12, H11-16.
Mindenes (burgonya): M1, M2, M3, M4, M5, M6, M7, M8, M11,
M12, M13, M17, M18, M20, M23, M24,
M28, M29, M32, M35, M36, M37.
Somogyi Kifli (burgonya): SK 1, SK 2, SK 4.
A negatív kontrollként használt nem transzgénikus dohánynövények
mindegyikén a fertőzést követő 7-10. napon megjelentek az első szisztemikus
tünetek. A levélér kivilágosodást, később érnekrózis és levélfoltosodás követte.
A nem transzgénikus kontroll- és a transzgénikus burgonyanövények között
tünetes növényt nem találtunk, sem vizuálisan, sem a dot-blot vizsgálatok által.
Az eredmények megerősítéséhez azonban a burgonyavonalakat szántóföldi
vizsgálatokban teszteltük tovább. A transzgénikus dohánynövények között sem
találtunk ebben az időszakban megbetegedett növényt. Néhány Virgin D
dohányvonal esetében (VD2, VD3, VD4, VD6) egy hónappal a fertőzés után
érnekrózis jelent meg, vonalanként 1 vagy 2 növénynél, de minden esetben az 5
µg/ml–es koncentrációval fertőzöttek között (VD2 –5A, 5C; VD3 –5B, 5C;
VD4 –5C; VD6 –5A, 5B) (2. táblázat és 13. ábra). Két hónappal a fertőzés után
a tünetes transzgénikus növények fiatal levelei tünetmentesekké váltak, ez a

67
„recovery” jelensége (Lindbo és Dougherty, 1992a; Lindbo és mtsai, 1993). A
vizsgált Virgin D vonalak közül 2-ben (VD1, VD5) nem találtunk
megfertőződött vagy „recovery” növényeket sem a vizuális vizsgálatok által,
sem a dot-blot eredmények alapján.
2. táblázat
Az üvegházi rezisztenciavizsgálat eredményei Virgin D növények esetében
Transzgénikus Virgin D dohányvonalak
VD1 VD2 VD3 VD4 VD5 VD6PVY konc.
(µg/ml)0.5 5 20 0.5 5 20 0.5 5 20 0.5 5 20 0.5 5 20 0.5 5 20
A E E E E R E E E E E E E E E E E R E
B E E E E E E E R E E E E E E E E R E
Növényegyedek C E E E E R E E R E E R E E E E E E E
Rövidítések: A, B, C –a növényegyedek megkülönböztető jelölése, E –Ellenálló, R –Recovery.

68
13. ábra
Transzgénikus dohánynövények összehasonlítása
a fertőzést követően 1 hónappal
Transzgénikus ellenálló (1.- VD1-5C) és megfertőződött, de későbbtünetmentessé váló- „recovery” (2.- VD6-5B) dohánynövények.
A Stamm C2 és a Hevesi 11 dohányfajták vizsgált transzgénikus vonalai
között nem találtunk megfertőződött növényeket sem a vizuális vizsgálatok
által, sem a dot-blot eredmények alapján. Ezt a tünetmentességet a növények
életük végéig megőrizték.
A dot-blot hibridizációs eljárás eredményeivel bizonyítottuk, hogy a
tünetmentes növények nem tartalmaztak vírust. Azok a növények, amelyeknél
érnekrózist, később „recovery” jelenséget tapasztaltunk, gyenge jelet adtak,
tehát kis mennyiségben, de akkumulálták a kórokozót, majd később ezek a
növények is vírusmentesekké váltak. A 14. ábrán a fertőzést követően egy
hónappal begyűjtött minták dot-blot analízisének részlete látható. Ekkor a

69
megfertőződött transzgénikus növények még vírustünetesek voltak és a vírust is
hordozták.
14. ábra
Dot-blot analízis
1. transzgénikus „recovery” dohánynövényből származó RNS kivonat, 2.rezisztens transzgénikus dohánynövény RNS kivonata, 3. megfertőződött, nemtranszgénikus kontrollnövény RNS kivonata, 4. vírus RNS hígítási sor.
4.2.4. A molekuláris vizsgálatok eredményeinek összegzése
Az előállított transzgénikus dohány- és burgonyavonalakkal végzett
molekuláris vizsgálatok eredményeinek összefoglalását tartalmazza a 3. és 4.
táblázat.
A Virgin D dohányfajta 38 vonalából mind a 38 termelte a köpenyfehérjét.
Az üvegházi körülmények között, mesterségesen fertőzött 6 vonalból 4 vonal 1-

70
2 növénye esetében tapasztaltunk „recovery” jelenséget, a többi ellenállónak
bizonyult. A Stamm C2 dohányfajta 55 előállított vonalából 37 termelte a
köpenyfehérjét, de 10 vonal esetében az expresszió alacsony szintű volt. A 12
fertőzött vonalból egyben sem találtunk vírusfertőzésre utaló jeleket sem
vizuálisan, sem a dot-blot vizsgálat eredményei alapján. A Hevesi 11
dohányfajta esetében 23 transzgénikus vonalat állítottunk elő, melyek közül 17
vonalban volt köpenyfehérje termelődés. A 12 fertőzött vonal mindegyike
vírus- és tünetmentesnek bizonyult a molekuláris és az üvegházi
vizsgálatokban.
Előállítottunk 37 Mindenes burgonyavonalat, melyek közül 34 esetben
tudtunk köpenyfehérje termelődést kimutatni. A Somogyi Kifli
burgonyafajtából 4 transzgénikus vonalat állítottunk elő és 3 esetben volt
kimutatható a köpenyfehérje növényen belüli termelődése.
3. táblázat
Transzgénikus dohányvonalak laboratóriumi- és vírusrezisztencia
vizsgálatainak összefoglalása.
DohányfajtákTranszgénikus
vonalak száma
Köpenyfehérjét
termelő vonalak
Vizsgált/
fertőzött vonal
Virgin D 38 38
6/4
(1-2 „recovery”
növ. /vonal)
Stamm C2 55 37 12/0
Hevesi 11 23 17 12/0

71
4. táblázat
Transzgénikus burgonyavonalak laboratóriumi vizsgálatainak összefoglalása
BurgonyafajtákTranszgénikus
vonalak száma
Köpenyfehérjét termelő
vonalak
Mindenes 37 34
Somogyi Kifli 4 3
4.2.5. Szántóföldi vizsgálatok
4.2.5.1. Dohányvonalak szabadföldi vizsgálata
Az üvegházi körülmények között ellenállónak bizonyult és a
rendelkezésünkre álló magok alapján, más kiválasztott vonalak közül néhányat
szántóföldi körülmények között vizsgáltuk tovább. Mindezt Dr. Nagy Gyula
közreműködésével, az AGROTAB Nemesítő és Vetőmagtermeltető Kft-vel
(Debrecen-Pallag) együttműködve végeztük. 19 Virgin D és 4 Stamm C2
transzgénikus vonal bevonását terveztük további vizsgálatokba.
Virgin D: VD1, VD2, VD3, VD4, VD5, VD6, VD8, VD9, VD10, VD11,
VD13, VD14, VD15, VD16, VD17, VD18, VD20, VD21, VD34.
Stamm C2: SC1, SC2, SC3, SC4.
Az 1998-as 27-es a Géntechnológiai tevékenységről szóló törvény
értelmében kért és kapott engedély mellett mindeddig 4 kiválasztott
transzgénikus vonal vehetett részt szántóföldi vizsgálatokban: Virgin D 5 és 17,

72
Stamm C2 1 és 4. A VD 5 és 17, illetve az SC 4 termelték a köpenyfehérjét,
míg az SC 1 nem. A kísérleti területre kiültetett vonalakban a természetes úton
történő vírusfertőzöttséget vizuálisan vizsgáltuk. A 2000-ben és 2001-ben
vonalanként 25-25 tő kiültetésére került sor. A PVY fertőzöttség mellett, mely
közepes erősségű volt mindkét évben, előfordultak más vírusok is, pl. CMV és
TMV. A transzgénikus növények ellenállónak bizonyultak a PVY fertőzéssel
szemben is (15. ábra), míg a fogékony fajtákban, mint pl. a Coker 254, 100%-os
PVY fertőzést tapasztaltunk (16. ábra). A további vizsgálatok elvégzése
érdekében a transzgénikus növények szántóföldi kiültetésére vonatkozó újabb
engedélyeztetésre kell várnunk.
15. ábra
Vírusellenálló transzgénikus dohánynövények szántóföldi kísérletben
(Debrecen-Pallag)

73
16. ábra
Az előtérben PVY-nal megfertőződött fogékony, nem transzgénikus
dohánynövények, a háttérben transzgénikus, vírusellenálló növények
(Debrecen-Pallag)
4.2.5.2. A transzgénikus burgonyavonalak szabadföldi vizsgálata
A transzgénikus burgonyavonalak szántóföldi vizsgálatokba történő
bevonására két szakaszban került sor. A kísérletek elsődleges célja a
vírusrezisztencia mesterséges illetve természetes körülmények közötti
megnyilvánulásának vizsgálata volt. A többi vizsgálat adatai főképpen
tájékoztató jellegűek, melyekkel a transzgénikus növények egyéb
tulajdonságainak változását illetve változatlanságát kívántuk figyelemmel
kísérni. Az 1999-ben vírusmentesített vonalak mechanikai fertőzése, ELISA
tesztje és a szabadföldi infektor növényes kísérlet 2000-ben, ugyanezen vonalak
kertészeti oltással történő vizsgálata, illetve a morfológiai-, gazdasági- és

74
felhasználhatósági értékvizsgálatai 2001-ben történtek. A 2000-ben
vírusmentesített vonalak mesterséges mechanikai fertőzésére PVYNTN és
PVYO/C izolátumokkal, majd ezek ELISA vizsgálatára és a Solanum demissum
A6 biotesztjére pedig 2001-ben került sor.
A transzgénikus burgonyavonalak vírus-ellenállóságának vizsgálata
PVYNTN fertőzést követő ELISA módszerrel történt (5. táblázat). Az
eredmények azt mutatják, hogy a transzgénikus burgonyavonalak nagy része
megfertőződött PVY-nal. Három vonal esetében azonban minden növény tünet-
és vírusmentes volt nemcsak vizuálisan, hanem az ELISA eredmények alapján
is. Ezek a Mindenes 11, Mindenes 12 és Mindenes 21 vonalak. A 37 illetve 40-
40 vizsgált növény egyike sem fertőződött meg a vírussal.

75
5. táblázat
Az 1999-ben vizsgálatokba vont transzgénikus burgonyavonalak (és kontrollok)
PVYNTN fertőzésének ELISA eredményei.
BurgonyavonalakPVY-nal megfertőződött növények/
összes növény
nem trg. Somogyi Kifli kontroll 40/40
Somogyi Kifli 2 37/37
nem trg. Mindenes kontroll 38/38
Mindenes 3 40/40
Mindenes 5 20/20
Mindenes 6 18/18
Mindenes 10 39/39
Mindenes 11 0/37
Mindenes 12 0/40
Mindenes 21 0/40
A kisparcellás, szabadföldi kísérlet elsődleges célja a rezisztencia illetve a
vírusfertőzés megjelenésének vizuális nyomon követése volt. A tüneti
felvételezés és az adatok többnyire előzetes, tájékoztató jellegűek. A vonalak
több ismétléses vizsgálata a következő évben folytatódik.
A kiültetett transzgénikus burgonya állomány vírusfertőzöttségének
vizuális vizsgálatakor begyűjtött adatok (6. táblázat) egyrészt azt mutatják,
hogy a növények mekkora %-a milyen erősségű tüneteket mutatott, azaz
táblázatban szereplő %-os adatok az adott vonalban a legsúlyosabb, feltüntetett
tüneterősségre vonatkoznak. A fennmaradó % érték az ennél enyhébb tüneteket

76
mutató növényegyedek arányát jelenti. Nyomon követhető a táblázatban az
egyes növényi vonalak magasságának alakulása, illetve kiegyenlítettsége is,
mely azt az értéket adja meg, hogy a növények mekkora %-a tartozott az
átlagmagassághoz.
6. táblázat
Transzgénikus burgonyavonalak vizuális felvételezése infektor növényes
szabadföldi kísérletben.
Kiültetett
vonalak
Vírusfertőzöttség
mértéke (PVY)
Magasság (átlag)
(cm)
Kiegyenlítettség
(%)fertőzött Somogyi
Kifli (SK)10% súlyos 60 90
SK-2 tünetmentes 55 80
SK-2 tünetmentes 55 80
fertőzött SK 10% súlyos 55 80
vírusmentes SK tünetmentes 50 80
vírusmentes SK tünetmentes 50 80
fertőzött SK 10% súlyos 50 80
Mindenes-3 90% súlyos 40-45 60
Mindenes-3 10% enyhe 40-45 60
fertőzött SK 15% súlyos 65 80
Mindenes-5 80% súlyos 35 50
Mindenes-6 95% súlyos 35 40
fertőzött SK 15% súlyos 55 80
Mindenes-10 80% súlyos 35 50
Mindenes-10 80% súlyos 35 50
fertőzött SK 20% súlyos 55 90
vírusmentes Mindenes 97,5% súlyos 40 60
vírusmentes Mindenes 97,5% súlyos 40 60

77
6. táblázat folytatása
Kiültetett
vonalak
Vírusfertőzöttség
mértéke (PVY)
Magasság (átlag)
(cm)
Kiegyenlítettség
(%)fertőzött SK 25% súlyos 55 80
Mindenes-11 tünetmentes 70 80
Mindenes-11 tünetmentes 70 80
fertőzött SK 20% súlyos 60 85
Mindenes-12 tünetmentes 75 80
Mindenes-21 tünetmentes 75 80
fertőzött SK 25% súlyos 55 80
Mindenes-21 tünetmentes 75 80
fertőzött SK 35% súlyos 40 55
Az eredmények alapján Mindenes 11, 12 és 21-es vonalak tünetmentesnek
bizonyultak. A táblázatban szereplő Somogyi Kifli 2 és SK vírusmentes
kontrollok meglepő, korábbi táblázatokban leírt eredményeinktől eltérő
adatokra a vírus hosszabb lappangási ideje adhat magyarázatot.
A szabadföldi kísérletekben kitűnő Mindenes 11, 12 és 21-es vonalak
vírus-ellenállóságának további tesztelése a következő évben kertészeti
oltásokkal folytatódott. PVYNTN-nel fertőzött dohány alanyra oltva a
burgonyavonalakat 2-2 növény esetében történt a víruskoncentráció
alakulásának nyomon követése, mely érték minden esetben a fertőzési határ
alatt mozgott. A fertőzött paradicsom alany esetében a víruskoncentráció nulla
vagy ahhoz közeli értékeket mutatott az oltóágban, míg az alanyban a
határértékek 10-14-szerese volt. Ez a kísérlet vonalaként 3-3 növény
bevonásával történt. Egészséges burgonya alanyra oltott fertőzött paradicsom

78
oltóág esetében is hasonlóan alakultak az eredmények. Míg a fertőzött
oltóágban a víruskoncentráció a határérték 10-14-szerese volt, addig a
transzgénikus burgonya alanyokban a vírus nem volt kimutatható. A
vonalanként vizsgált 3-3 növény mindegyike vírusmentes volt.
Morfológiai-, gazdasági- és felhasználási értékvizsgálatok elvégzésére is sor
került (7. táblázat). Az áprilisi ültetést követően a kelési % meghatározása 10-
15 cm-es méret elérésekor következett. Volt olyan növény, amely ebben az
időpontban még nem kelt ki (Mindenes 10), de a második felvételezéskor
(június közepén) már volt értékelhető adat. Az átlagos növénymagasság
felvételezése ekkor történt. A hektáronkénti termésmennyiség a
parcellamérethez viszonyítva lett kiszámolva, így a táblázatban szereplő adatok
közelítő értékeket adnak. A három, korábban már kiemelt Mindenes vonal (11,
12 és 21) nemcsak vírus-ellenállóságával tűnt ki a többi transzgénikus
burgonyavonal közül, hanem a termésmennyiségük is átlagon felüli volt. A
keményítő %, mely fajta-függő, transzgénikus vonalainkban is a magyarországi
17.5%-os átlagérték körül mozgott. A már említett három Mindenes vonal a
keményítő % alakulása szerint az alábbi besorolás alapján a vegyesen
felhasználható burgonyák csoportjába sorolható:
A: sütésre való ill. salátaburgonya (13 -14%)
B: vegyesen felhasználható (15 -17 %)
C: burgonyapüré (18 -19%)
D: takarmány, szesz, ipari burgonya (19 -20% fölött)

79
7. táblázat
Transzgénikus burgonyavonalak kelése, fejlődése, termése és beltartalmi értéke.
Vonal
megnevezése Kelési %
Átl. növény-
magasság
(cm)
Termés
(t/ha)
Keményítő
%
Somogyi K. 2 90 55 25.0 17.61
SK kontroll 100 50 28.8 16.95
Mindenes 3 70 45 12.5 -
Mindenes 5 25 35 9.94 -
Mindenes 6 60 35 12.74 -
Mindenes 10 0 35 16.7 -
Mindenes 11 90 70 38.25 15.46
Mindenes 12 70 80 26.9 15.66
Mindenes 21 100 85 43.0 16.12
A 2001-ben szántóföldi vizsgálatokba bevont transzgénikus
burgonyavonalak (vonalanként 4-4 növényt) mesterséges fertőzése PVYNTN és
PVYO/C izolátumokkal történt. A fertőzést követő 37. napon került sor a DAS-
ELISA vizsgálatokra (8. táblázat). Negatív kontrollként nem transzgénikus in
vitro növények illetve a transzgénikus vonalak kiültetett, nem fertőzött egyedei
szerepeltek, pozitív kontrollként pedig mesterségesen megfertőzött Nicotiana
tabacum Xanti-nc, Mindenes és Somogyi Kifli növények.

80
8. táblázat
A 2001-ben szántóföldi tesztelésbe vont burgonyavonalak mesterséges
mechanikai fertőzésének ELISA-vizsgálati eredményei.
Fertőzött / vizsgált mintákVonalak megnevezése
PVYO/C PVYNTN
Mindenes (nem trg.) 1/1 1/1
Mindenes- 1 4/4 4/4
Mindenes- 3 2/4 4/4
Mindenes- 5 4/4 3/4
Mindenes- 6 3/4 4/4
Mindenes- 8 1/4 0/4
Mindenes- 9 0/4 0/4
Mindenes- 10 4/4 4/4
Mindenes- 14 3/4 4/4
Mindenes- 15 0/4 0/4
Mindenes- 16 0/4 4/4
Mindenes- 17 4/4 4/4
Mindenes- 18 2/2 3/3
Mindenes- 20 2/4 4/4
Mindenes- 27 4/4 4/4
Mindenes- 33 3/4 4/4
Somogyi Kifli (nem trg.) 1/2 1/2
Somogyi Kifli- 4 1/4 4/4
N. tabacum Xanthi-nc (nem trg) 2/2 2/2
A 9. és 10. táblázat a növényeken (vonalanként 4-4) okozott tüneteket
részletezi külön-külön a PVYNTN és PVYO/C izolátumok esetében. A táblázatok
tartalmazzák a Solanum demissum A6 teszt eredményeit is, azaz, hogy a

81
fertőzött transzgénikus növény tartalmaz-e vírust, illetve az átvihető-e a
tesztnövény levelekre. A vizsgálatokban három Mindenes vonal ( M8, M9 és
M15) eredményei tűnnek ki a többi közül. Ezek a vonalak eddigi adataink
alapján a mesterséges PVY fertőzéssel szemben ellenállónak bizonyultak.
9. táblázat
A PVYO/C törzzsel fertőzött transzgénikus növények tünetei és a S. demissum
A6 teszt eredményei
PVYO/C által okozott tünetek a tesztnövényekenTrg.
vonal
neve1. 2. 3. 4.
S. demissum
A6
M 1Mo, Nsp,
Lab
Mo, Nsp,
Nri, Vn
Mo, Nri,
Vn, Lab
Mo, Vn,
Lab+
M 3Nsp, Nri,
Cl, Vn
Nsp, Vn,
Clsp Neg.
Nsp,
Clsp+
M 5Nsp,
Vn
Nsp, Nri,
Vn, Lab
Nsp, Nri,
Clri, Lab
Nsp, Nri,
Vn+
M 6Nsp,
Vn Neg.
Vn,
Nri
Mo, Nsp,
Nri, Vn+
M 8Nsp, Nri,
Clsp, Vn Neg. Neg. Neg.+ (1 növény)
M 9Neg. Neg. Neg. Neg.
-
M 10Mo, Nsp,
Vn, Clsp
Nsp, Vn,
Stn, Lab
Nsp, Nri,
Vn, Stn
Mo, Vn,
Nsp, Nri+
M 14Nri,
Vn
Nsp, Nri,
Vn, Stn
Nsp, Vn,
Lab
Vn,
Lab+

82
9. táblázat folytatása
PVYO/C által okozott tünetek a tesztnövényekenTrg.
vonal
neve1. 2. 3. 4.
S. demissum
A6
M 15Neg. Neg. Neg. Neg.
-
M 16Vn, Clsp,
Lab Neg.
Mo, Nri,
Vn, Lab
Nsp, Vn,
Stn, Lab+
M 17Mo, Nsp,
Nri, Vn
Mo, Nsp,
Nri, Vn
Mo, Nsp,
Nri, Vn
Mo, Nsp,
Nri, Vn+
M 18Mo, Nsp,
Vn, Lab
Mo, Nsp,
Vn, Lab- - +
M 27Nsp, Nri,
Vn, Lab
Mo, Nsp,
Nri, Vn
Mo,
Vn
Mo, Nsp,
Nri, Vn+
M 33 Nsp Nsp, VnNsp, Vn,
Nri
Nsp, Vn,
Nri+
SK 4Nsp,
Lab
Nsp, Vn
LabNsp
Nsp, Vn
Lab+
SK kontr. Nsp Nsp - - +
M kontr. sMo, Vn sMo, Vn sMo, Vn sMo, Vn +Rövidítések: M- Mindenes, SK- Somogyi Kifli, Cl- klorózis, Clri- klorotikus gyűrű, Clsp-
klorotikus folt, Lab- levélhullás, Mo- mozaik, sMo- súlyos mozaik, Neg- negatív, Nri-
nekrotikus gyűrű, Nsp- nekrotikus folt, Stn- törpülés, Vn- érnekrózis.

83
10. táblázat
A PVYNTN törzzsel fertőzött növények tünetei és a S. demissum A6 teszt
eredményei
PVYNTN által okozott tünetek a növényekenTrg.
vonal
neve1. 2. 3. 4.
S. demissum
A6
M 1 mMo, Vn mMo, Vn Vn mMo, Vn +
M 3 Vn Vn Vn Vn +
M 5 sMo, Ld, Vn Vn Nsp Nsp +
M 6 Vn Vn Vn mMo +
M 8 Neg. Neg. Neg. Neg. -
M 9 Neg. Neg. Neg. Neg. -
M 10 Vn mMo, Vn mMo, Vn mMo, Vn, Ld +
M 14 Vn Vn Vn Vn +
M 15 Neg. Neg. Neg. Neg. -
M 16 mMo, Vn Vn sMo, Vn mMo, Vn +
M 17 Vn Lc, sMo, Vn Vn Vn +
M 18 mMo Lc, sMo, Stn Lc, sMo, - +
M 27 Vn Nsp Vn mMo, Vn +
M 33 Vn mMo, Vn mMo, Vn mMo, Vn +
SK 4 Vn Vn Vn Vn +
SK kontr. mMo Nsp - - +
M kontr. Mo, Ld, Vn Mo, Ld, Vn Mo, Ld, Vn Mo, Ld, Vn +Rövidítések: M- Mindenes, SK- Somogyi Kifli, Lc- levélgöndörödés, Ld- levéldeformáció,
Mo- mozaik, mMo- enyhe mozaik, sMo- súlyos mozaik, Neg- negatív, Nsp- nekrotikus folt,
Stn- törpülés, Vn- érnekrózis.

84
Az összes, korábban felsorolt transzformáns burgonyavonalat vizsgáltuk
előbb molekuláris vizsgálatokkal, majd többféle módszerrel szántóföldi
körülmények között is. A kísérleteink adatai azt mutatják, hogy a különböző
transzgénikus vonalakban a rezisztencia megnyilvánulása változó volt,
fogékony vonalak mellett gyenge és erős mértékű vírus-ellenállóságot mutató
növényeket is megfigyeltünk. A vizuálisan tünetmentes növényegyedek az
ELISA-vizsgálatok alapján is rezisztensnek bizonyultak. Két egymást követő
évben három olyan különböző Mindenes vonalat találtunk (M11, M12 és M21),
amelyek magas fokú rezisztenciával rendelkeztek a mesterséges, illetve
természetes körülmények között fertőző PVYNTN törzzsel. Ezek a vonalak más
vizsgálatokban is értékesnek bizonyultak, különösen a Mindenes 21, mely 43
t/ha termésmennyiséget adott. Azok közül a vonalak közül, melyek vizsgálatát a
2000-es évben kezdtük meg szintén három Mindenes vonal tűnt ki (M8, M9 és
M15), melyek fertőzést követő ELISA vizsgálatainak eredményei alapján
mesterséges PVY inokuláció esetén ellenállónak bizonyultak. A Mindenes 8-as
vonal vizsgált nyolc növénye közül egy azonban fertőződött PVYO/C-vel, és ezt
a Solanum demissum bioteszt is igazolta, a maradék 7 vizsgált M8-as növény
tünetmentessége azonban a vonal ellenállóképességének vizsgálatához további
kísérletek elvégzését teszi indokolttá. Így tehát az M8 vonal további 7 növénye
és az említett másik két vonal (M9 és M15) egyik növénye sem fertőződött meg
sem PVYNTN-nel, sem PVYO/C-vel. Ezeken a vizsgált növényeken nem találtunk
vírustüneteket és a vírus replikáció hiányát igazolták a Solanum demissum A6
bioteszt eredményei is.

85
5. EREDMÉNYEK MEGVITATÁSA, KÖVETKEZTETÉSEK
Az előállított transzgénikus vonalak közül a Virgin D dohányfajta esetében
38 vonalat hoztunk létre, melyek mindegyike hordozta a transzgént és magát a
köpenyfehérjét is termelte. Üvegházi kísérletben 6 vonalat fertőztünk
mesterségesen, tisztított PVY-H-val. Ezek a VD1, VD2, VD3, VD4, VD5, VD6
vonalak voltak. A VD1 és a VD5 vonalban nem találtunk fertőzött növényeket
sem vizuálisan, sem a dot-blot vizsgálatok által. A többi négy vonalban (VD2,
VD3, VD4, VD6) a fertőzés után egy hónappal érnekrózis megjelenését
figyeltük meg. A vírus akkumulációt a dot-blot vizsgálatok adatai is
alátámasztották. Minden esetben olyan növényeken tapasztaltuk ezt, melyeket
5µg/ml víruskoncentrációval fertőztünk. A fertőzést követő két hónap elteltével
azonban ezek a tünetek eltűntek, a növények újból vírusmentesek voltak,
melyet a vizuális megfigyelések mellett a dot-blot eredmények is igazoltak.
Ezeknél a vonalaknál a „recovery” jelenséget fedezhettük fel, azaz a kezdeti
fertőzést követően a növények fiatal levelei már vírusmentesen fejlődtek. A
kezdetben kialakuló tünetek enyhébbek voltak és késéssel jelentek meg a
kontrollokhoz képest, amelyeken már a fertőzést követő 7-10. napon
észrevehető és kimutatható vírusfertőzés tüneteit tapasztaltuk. Szántóföldi
körülmények között mindeddig két vonal (VD5 és VD17) vizsgálatára volt
lehetőségünk. A két éves vizsgálati idő alatt a PVY fertőzöttség közepes
erősségű volt. A fogékony fajták 100%-osan megfertőződtek, míg
transzgénikus vonalaink egészségesek maradtak.
A Stamm C2 dohányfajta esetében 55 transzgénikus vonalat hoztunk létre,
melyek mindegyikét vizsgáltuk molekuláris módszerekkel. A transzgén
terméket összesen 34 esetben tudtuk biztosan kimutatni, ebből 8 esetben az
expresszió alacsony szintű volt. Az üvegházi körülmények között,

86
mesterségesen megfertőzött 12 vonal mindegyike ellenállónak bizonyult mind
vizuálisan, mind a dot-blot vizsgálatok eredményei szerint. A 12 vonal között
voltak olyan vonalak, melyek alacsony szinten expresszálták a köpenyfehérjét
(SC35, SC46, SC47, SC48) és voltak olyanok, melyekből nem tudtuk kimutatni
a transzgén expressziót (SC36, SC39, SC40, SC41, SC43, SC44, SC45, SC49).
Ezzel párhuzamosan szántóföldi vizsgálatok elvégzésére 2 vonal esetében volt
lehetőségünk (SC1, SC4). Az SC1 nem termelte a köpenyfehérjét, míg az SC4
igen. A VD vonalaknál leírtak igazak az SC vonalak eredményeire is, azaz a
közepes erősségű PVY fertőzés mellett mindkét vizsgált transzgénikus vonal
ellenálló volt a vírussal szemben, ellentétben a fogékony, 100%-ban
megfertőződött fajtákkal.
A Hevesi 11 dohányfajtából 23 transzgénikus vonalat állítottunk elő, ezek
közül 17 vonal termelte a köpenyfehérjét. Az üvegházi vizsgálatokban 12 vonal
vett részt, melyek egy része nem termelte a CP-t (H11-12, H11-16), míg nagy
részük termelte (H11-1, H11-2, H11-3, H11-4, H11-6, H11-7, H11-8, H11-9,
H11-10, H11-11). A növények egyike sem fertőződött meg a tisztított PVY-H-
val történő inokulálást követően, amit a vizuális- és a dot-blot vizsgálatok is
igazoltak. Szántóföldi vizsgálatokban ezek a vonalak mindeddig nem vettek
részt.
A Mindenes burgonya fajtából 37 transzgénikus vonalat állítottunk elő,
melyek közül 34-ben tudtuk a köpenyfehérje termelődést kimutatni. A Somogyi
Kifli fajtából 4 transzgénikus vonalat kaptunk, ezek közül 3 esetben volt
köpenyfehérje termelődés. Az üvegházi körülmények között fertőzött 22
Mindenes és 3 Somogyi Kifli vonal egyike sem fertőződött a PVY-nal. Ezeket
az eredményeket szántóföldi vizsgálatokkal pontosítottuk. A szabadföldi
vizsgálatokban tesztelt 22 Mindenes és 4 Somogyi Kifli közül voltak olyan
vonalak, melyek magas szinten expresszálták a köpenyfehérjét (SK1, SK2, M1,

87
M3, M4, M7, M11, M12, M14, M15, M17, M18, M20, M21, M24, M27, M33),
néhány vonal esetében alacsony szintű CP termelődést tapasztaltunk (SK3, M2,
M5, M8, M10) és néhány vonal egyáltalán nem termelte a CP-t (SK4, M6, M9,
M16). A két éven át tartó különféle szántóföldi vizsgálatokban folyamatosan
ugyanazok a vonalak tűntek ki ellenállóságukkal: Mindenes 11, Mindenes 12 és
Mindenes 21. Mindhárom vonalban tapasztaltunk transzgén eredetű
köpenyfehérje termelődést. A transzgénikus vonalak második csoportjával
végzett kísérletek alapján szintén három vonal tűnt ki: Mindenes 8, Mindenes 9
és Mindenes 15. Az M8-ban alacsony szintű volt a kimutatható CP, az M9 nem
termelte, az M15 pedig termelte a CP-t.
A vírussal szemben ellenálló vonalak között tehát CP-t termelő-, nem
termelő- és alacsony szinten termelő vonalak is előfordultak Ezek alapján azt a
következtetést vontuk le, hogy a PVY-nal szembeni rezisztencia, a korábbi, más
kutatócsoportok által leírt eredményekhez hasonlóan, nem fehérjetermék-függő.
Ez a Potyvírusok mindegyikére általánosan is jellemző. Regner és mtsai (1992)
például PPV CP génnel transzgénikus N. benthamiana és N. clevelandii
növényeknél tapasztalták, hogy a PPV-vel szembeni ellenállóság kiváltásához
nem szükséges a fehérjetermék jelenléte. Lindbo és Dougherty (1992b) szintén
RNS-függő rezisztenciát tapasztaltak TEV CP gént transzgénként hordozó
dohánynövények TEV-vel történő fertőzése esetén. Számos más kísérlet is
bizonyítja, alátámasztva az eredményeinket, hogy a Potyvírusoknál nincs
összefüggés a rezisztencia mértéke és a transzgén expressziós szintje között
(van der Vlugt és mtsai, 1992, Kollár és mtsai, 1993, Smith és mtsai, 1995,
Hammond és Kamo, 1995).
A munkánk során tapasztaltak alapján elmondható, hogy a védettséget
elsősorban maga a transzgén vagy az erről átíródott transzkriptum (RNS) váltja
ki. Az ellenállóság kialakulását nagy valószínűség szerint a géncsendesítés

88
jelensége idézi elő, mely jellemzően alacsony vagy nem detektálható transzgén
expresszióval jár együtt. Az üvegházi vizsgálatoknál tapasztalt „recovery”
jelenség magyarázata is a géncsendesítés lehet. Ez a szekvenciaspecifikus
lebontó mechanizmus transzgénikus növényeknél a „küszöbérték modell”
(Smith és mtsai, 1994) szerint úgy nyilvánulhat meg, hogy a nagy
mennyiségben jelenlévő, transzgén eredetű RNS-ek mennyisége meghalad egy
szintet, mely beindítja a citoplazmatikus RNS degradáló folyamatot. Ennek
következtében a transzgénnel homológ fertőző vírus-eredetű nukleinsavak is
lebontódnak, ezáltal elmarad a vírusfertőzés és a növények ellenállóak lesznek a
későbbi vírustámadásokkal szemben is. Goodwin és mtsai (1996) szerint a
„recovery” jelenség magyarázatához tartozhat a transzgén alacsonyabb, egy-két
kópiás beépülése a genomba, míg a rezisztencia kialakulásához három vagy
több kópia szükséges. A „recovery” jelenség esetében a transzgén eredetű RNS
mennyiség a vírus eredetű transzkriptumokkal együtt éri el a határértéket, mely
beindítja a specifikus nukleinsav lebontást, így a kezdeti fertőzést vírusmentes
állapot követi. A géncsendesítés beindításával, a szignálmolekula terjedésével
és a specifikus nukleázok kialakulásával a növény egész élettartamára megőrzi
ellenállóképességét a transzgénnel, illetve transzkriptumával homológ
nukleinsavval rendelkező vírusokkal szemben.
A transzgénikus növények vizsgálatai alatt nem tapasztaltuk semmiféle
hátrányos tulajdonság megjelenését. A transzgénikus burgonyavonalak
beltartalmi értéke az átlag körül mozogott, míg egyes vonalak terméshozama a
Magyarországon átlagosnak ítélt 17-19 t/ha-hoz képest jelentősen nagyobb volt:
38, 26 és 43 t/ha. Ezeket a kiugró értékeket a termésmennyiség tekintetében a
már említett Mindenes 11, a Mindenes 12 és a Mindenes 21 transzgénikus
vonalak esetében tapasztaltuk.

89
Eredményeink azt mutatják, hogy a vírus eredetű köpenyfehérje gén
növényekbe való beépítése és ezáltal ellenálló fajták létrehozása haszonnal
járulhat a növénynemesítési munkákhoz. Hasonlóan a külföldi eredményekhez,
ahol más burgonyafajták esetében pl. Russet Burbank, Bintje már létrehoztak
PVY-nal szemben ellenálló transzgénikus burgonyavonalakat, hazánkban is
fontossá vált az itt előforduló PVY vírusizolátum felhasználásával hazai fajták
vírusellenállóvá alakítása. Munkánk így a magyarországi növénytermesztés,
elsősorban a burgonyatermesztés számára jelentős lépés, mivel a
köztermesztésből kiszorult régi magyar fajták - melyek mára már többnyire
csak génbankokban találhatóak meg - újra visszakerülhetnek a szántóföldekre,
bővítve a mezőgazdaság számára hasznosítható növényeink körét.

90
6. ÖSSZEFOGLALÁS
A vírusok elleni védekezésben az egyik leghatékonyabb megoldás
vírusrezisztens transzgénikus növények előállítása. Az elmúlt két évtizedben a
molekuláris biológiai eszközök felhasználásával a világ különböző
laboratóriumaiban vírusellenálló növényeket állítottak elő. Az általunk is
alkalmazott módszer a köpenyfehérje gén által indukált keresztvédettségi
reakción alapul. A növénytermesztésben nagy károkat okozó burgonya Y
vírussal (PVY) szembeni ellenállóságot váltottunk ki transzgénikus növények
létrehozásával. A PVY-H (magyar izolátum) köpenyfehérje (CP) génjét
Agrobacterium tumefaciens vektor segítségével építettük burgonya- és
dohánynövényekbe. A transzformált sejteket szelektív táptalajon illetve
üvegházban egész növényekké neveltük, majd molekuláris módszerekkel
vizsgáltuk. A kísérlethez 3 dohányfajtát használtunk: Virgin D, Stamm C2,
Hevesi 11, melyekből sorrendben 38, 55 illetve 23 transzgénikus vonalat
sikerült előállítani, valamint 2 burgonyafajtát: Mindenes és Somogyi Kifli,
melyekből 37 és 4 vonalat állítottunk elő. A transzgén integrációt PCR technika
segítségével ellenőriztük, a transzkripciót illetve transzlációt Northern és
Western analízisekkel mutattuk ki. A vírus-ellenállóságot üvegházi
körülmények között mesterségesen, tisztított PVY-H-val történő fertőzést
követő dot-blot analízissel vizsgáltuk. A kiválasztott vonalak szántóföldi
tesztelésre kerültek. A természetes körülmények között fertőző vírussal
szemben a négy vizsgált dohányvonal (VD 5 és 17, SC 1 és 4) ellenállónak
bizonyult. A szántóföldi vizsgálatok eredményei által hat Mindenes vonalat
találtunk, melyek nagy fokú ellenállóságot mutattak a vírussal szemben (M 8, 9,
11, 12, 15, 21). Eredményeink rávilágítanak arra, hogy ez a módszer nagy
hatékonysággal és biztonsággal használható a szántóföldi növénytermesztésben.

91
ABSTRACT
To protect the cultivated plants from virus diseases one of the most
effective solution is to produce transgenic plants. During the last twenty years
virus resistant transgenic plants were produced by genetic engineering in
several laboratories all over the world. The method used by us is based on the
cross-protection phenomenon caused by the coat protein. We produced resistant
transgenic plants against PVY (potato virus Y), which causes severe loss in
crop production. The CP (coat protein) gene of the PVY-H (hungarian isolate)
was used by the help of Agrobacterium tumefaciens binary vector, to produce
tobacco and potato transformants. The plantlets were grown on selective media
and later in greenhouse conditions. We used three tobacco cultivars: Virgin D,
Stamm C2 and Hevesi 11, and we got 38, 55 and 23 transgenic lines. The two
potato cultivars are: Mindenes and Somogyi Kifli, and we got 37 and 4
transgenic lines. The integration of the coat protein gene to the plant was
confirmed by polymerase chain reaction, the transcription and the translation
were followed by Northern and Western blots. To test the resistance against
viruses we infected the plants in greenhouse with purified PVY-H and made
dot-blot tests. Some of the lines were choosen and grown in field conditions.
The four tested tobacco lines (VD 5 and 17, SC 1 and 4) were resistant against
the naturally infecting PVY, and we found six Mindenes lines (M 8, 9, 11, 12,
15, 21) which also shown high level of resistance.
Our results show that this method is safe and effective, and give new
possibilities for the plant breeding but especially for old hungarian potato
cultivars to get back to the fields and to save the biodiversity of the agriculture.

92
7. IRODALOM
Allison, R., Johnson, R. E. and Dougherty, W. G. (1986): The nucleotide sequence of the codingregion of tobacco etch virus genomic RNA: evidence for the synthesis of a single polyprotein.Virology 154: 9-20.
Anderson, E. J., Stark, D. M., Nelson, R. S., Beachy, R. N. (1989): Transgenic plants thatexpress the coat protein genes of tobacco mosaic virus of alfalfa mosaic virus interfere withdisease development of some nonrelated viruses. Phytopathology 79: 1284-1290.
Angenent, G. C., van den Ouveland, J. M. and Bol, J. F. (1990): Susceptibility to virus infectionof transgenic tobacco plants expressing structural and nonstructural genes of tobacco rattlevirus. Virology 175: 191-198.
Ares, X., Calamante, G., Cabral, S., Lodge, J., Hemenway, P., Beachy, R. N. and Mentabery, A.(1998): Transgenic plants expressing potato virus X ORF 2 protein (p24) are resistant totobacco mosaic virus and Ob tobmoviruses. Journal of Virology 72: 731-738.
Atreya, P. L., Atreya, C. D. and Pirone, T. P. (1991): Amino acid substitutions in the coatprotein results in loss of insect transmissibility of a plant virus. Proc. Natl. Acad. Sci. USA 88:7887-7891.
Atreya, C. D. (1992): Application of genome sequence information in potyvirus taxonomy: anoverview. In: Barnett, O. W. (ed.) Potyvirus taxonomy. Springer, New York 1992.
Balázs, E. és Gáborjányi, R. (1984): A génátültetés alkalmazása: a korszerű növényvédelem újlehetősége. Növényvédelem 20: 145-151.
Balázs, E. (1997): A növényvédelem új feladata a géntechnológiai úton módosítottszervezetekről alkotott törvény végrehajtása. Növényvédelem 33: 582-584.
Barker, H., Reavy, B., Kumar, A., Webster, K. D., Mayo, M. A. (1992): Restricted virusmultipication in potatoes transformed with the coat protein gene of potato leafroll luteovirus:similarities with a type of host gene-mediated resistance. Ann. Appl. Biol. 120: 55-64.
Bass, B. L. (2000): Double stranded RNA as a template for gene silencing. Cell 101:235-238.
Baulcombe, D. C. (1996): Mechanism of pathogen-derived resistance to viruses in transgenicplants. The Plant Cell 8: 1833-1844.
Baulcombe, D. C. and English, J. J. (1996): Ectopic pairing of homologous DNS and post-transcriptional gene silencing in transgenic plants. Current Opinion in Biotechnology 7: 173-180.
Beachy, R. N. (1990): Coat protein-mediated resistance against virus infection. Annu. Rev.Phytopathol. 28: 451-474.

93
Beczner, L., Horváth, J., Romhányi, I., Förster, H. (1984): Studies on the etiology of tubernecrotic ringspot disease in potato. Potato Res. 27: 339-352.
Bendahmane, M., Fitchen, J. H., Zhang, G. and Beachy, R. N. (1997): Studies of coat protein-mediated resistance to tobacco mosaic tobamovirus: correlation between assembly of mutantcoat proteins and resistance. Journal of Virology 71: 7942-7950.
Bertioli, D. J., Cooper, J. I., Edwards, M. L., Hawes, W. S. (1992): Arabis mosaic nepoviruscoat protein in transgenic tobacco lessens disease severity and virus replication. Ann. Appl.Biol. 120: 47-54.
Bevan, M. W., Mason, S. E., Goelet, P. (1985): Expression of tobacco mosaic virus coat proteinby a cauliflower mosaic virus promoter in plants transformed by Agrobacterium. EMBO J.4:1921-1926.
de Bokx, J. A. and Huttinga, H. (1981): Potato Virus Y. CMI/AAB Description of Plant VirusesNo. 242.
Bourdin, D. and Lecoq, H. (1991): Evidence that heteroencapsidation between two potyvirusesis involved in aphid transmission of a non-aphid-transmissible isolate from mixed infections.Phytopathology 11: 1459-1464.
Brakke, M. K., Ball, E. M., Hsu, Y. H. and Langenberg, W. G. (1987): Wheat streak mosaicvirus cylindrical inclusion body protein. Journal of General Virology 68: 281-287.
Brault, V., Candresse, T., le Gall, O., Delbos, R. P., Lanneau, M. and Dunez, J. (1993):Genetically egineered resistance against grapevine chrome mosaic nepovirus. Plant Mol Biol.21: 89-97.
Brigneti, G., Voinnet, O., Li, W. X., Ji, L. H., Ding, S. W. and Baulcombe, D. C. (1998): Viralpathogenicity determinants are suppressors of transgene silencing in Nicotiana benthamiana.EMBO J. 17: 6739-6746.
Bruyere, A., Wantoba, M., Flasinski, S., Dzianott, A. and Bujarski, J. J. (2000): Frequenthomologous recombination events between molecules of one RNA component in a multipartiteRNA virus. J. of Virology, 9: 4214-4219.
Burgyán J., Perkáta, K., Salamon P., Havelda, Z. (1998): A paradicsom foltoshervadás-vírushazai izolátumának molekuláris jellemzése és vírusrezisztens dohány előállítása genetikaitranszformációval. Növényvédelem 11:607-612.
Candelier-Harvey, P. and Hull, R. (1993): Cucumber mosaic virus genome is encapsidated inalfalfa mosaic virus coat protein expressed in transgenic tobacco plants. Transgenic Research 2:277-285.
Carrington, J. C. and Dougherty, W. G. (1987): Small nuclear inclusion protein encoded by aplant potyvirus genome is a protease. Journal of Virology 61: 2540-2548.

94
Carrington, J. C., Freed, D. D. and Sanders, T. C. (1989): Autocatalytic processing of thepotyvirus helper component proteinase in Escherichia coliand in vitro. Journal of Virology 63:4459-4463.
Carrington, J. C., Jensen, P. E. and Schaad, M. C. (1998): Genetic evidence for an essentialrole for potyvirus CI protein in cell-to-cell movement. Plant Journal 14: 393-400.
Carrington, J. C., Kasschau, K. D. and Johansen L. K. (2001): Activation and suppression ofRNA silencing by plant viruses. Virology 281: 1-5.
Cassidy, B. G. and Nelson, R. S. (1995): Differences in protection phenotypes in tobacco plantsexpressing coat protein genes from peanut stripe potyvirus with or without an engineered ATG.Mol. Plant-Microbe Interact. 8: 357-365.
Chachulska, A. M. (1998): Potato virus Y phylogeny and engineered virus resistance. Ph.D.Thesis, Warsaw 1998. pp 106.
Chia, T. F., Chan, Y. S. and Chua, N. H. (1992): Characterization of cymbidium mosaic viruscoat protein gene and its expression in transgenic tobacco plants. Plant Mol. Biol. 18: 1091-1099.
Chiang, C-H., Wang, J-J., Jan, F-J., Yeh, S-D. and Gonsalves, D. (2001): Comparativereactions of recombinant papaya ringspot viruses with chimeric coat protein (CP) genes andwild-type viruses on CP-transgenic papaya. J. of General Virology 82:2827-2836.
Chowrira, G. M., Cavileer, T. D., Gupta, S. K., Lurquin, P. F., Berger, P. H. (1998): Coatprotein-mediated resistance to pea enation mosaic virus in transgenic Pisum sativum L.Transgenic Res. 7: 265-271.
Clark, M. F. and Adams, A. N. (1977): Characteristics of the microplate method of enzyme-linked immunosorbent assay for the detection of plant. J. Gen. Virol. 34: 475-483.
Clark, W. G., Register, J. C., Nejidat, A., Eichholtz, D. A., Sanders, P. R., Fraley, R. T. andBeachy, R. N. (1990): Tissue-specific expression of the TMV coat protein in transgenic tobaccoplants affects the level of the coat protein-mediated virus protection. Virology 179: 001-008.
Clark, W. G., Fitchen, J., Nejidat, A., Deom, C. M. and Beachy, R. N. (1995): Studies of coatprotein-mediated resistance to tobacco mosaic virus (TMV). II. Challenge by a mutant withaltered virion surface does not overcome resistance conferred by TMV coat protein. J. ofGeneral Virology 76: 2613-2617.
Covey, S. N., Al-Kaff, N. S., Lángara, A. and Turner, D. S. (1997): Plants combat infection bygene silencing. Nature 385: 781-782.
Cuozzo, M., O'Conell, K. M., Kaniewski, W., Fang, R. X., Chua, N. H. and Tumer, N. E. (1988):Viral protection in transgenic tobacco plants expressing the cucumber mosaic virus coat proteinor its antisense RNA. Bio/Technology 6: 549-555.

95
Da Camara Machado, M. L., da Camara Machado, A., Mattanovich, D., Regner, F., Hanzer,V., Durniok, B., Steinkeller, H., Himmler, G., Katinger, H. (1990): Expression of the plum poxcoat protein in Nicotiana clevelandii. Acta Horticulturae 280: 577-580.
Da Camara Machado, L. M., da Camara Machado, A., Hanzer, V., Weiss, H., Regner, F.,Steinkeller, H., Mattanovich, D., Plail, R., Knapp, E., Kalthoff, B., Katinger, H. (1992):Regeneration of transgenic plants of Prunus armeniaca containing the coat protein gene ofPlum pox virus. Plant Cell Reports 11: 25-29.
Dalmay, T., Hamilton, A., Mueller, E. and Baulcombe, D. (2000a): Potato virus X amplicons inArabidopsis mediate genetic and epigenetic gene silencing. The Plant Cell, 12: 369-379.
Dalmay, T., Hamilton, A., Rudd, S., Angell, S. and Baulcombe, D. (2000b): An RNA-dependentRNA polymerase gene in Arabidopsis is required for posttranscriptional gene silencingmediated by a transgene but not by a virus. Cell, 101: 543-553.
Dinant, S., Balise, F., Kuziak, C., Astier-Manifacier, S. and Albouy, J. (1993): Heterologousresistance to potato virus Y in transgenic tobacco plants expressing the coat protein gene oflettuce mosaic virus. Phytopathology 83: 818-824.
Dolja, V. V. and Carrington, J. C. (1992): Evolution of positive-strand RNA viruses. Semin.Virol. 3: 315-326.
Dolja, V. V., Haldeman- Cahill, R., Montgomery, A. E., Vadenbosch, K. A., Carrington, J. C.(1995): Capsid protein determinants involved cell-to-cell and long distance movement oftobacco etch potyvirus. Virology 206: 1007-1016.
Dougherty, W. G. and Carrington, J. C. (1988): Expression and function of potyviral geneproducts Ann. Rev. Phytopathology 26: 123-143.
Dougherty, W. G., Lindbo, J. A., Smith, H. A., Parks, T. D., Swaney, S. and Proebsting, W. M.(1994): RNA-mediated virus resistance in transgenic plants: exploitation of a cellular pathwaypossibly involved in RNA degradation. Mol. Plant-Microbe. Interact. 7: 544-552.
van Dun, C. M. P., Bol, J. F. and van Vioten-Doting, L. (1987): Expression of alfalfa mosaicvirus and tobacco rattle virus coat protein genes in transgenic tobacco plants. Virology 159:229-305.
van Dun, C. M. P. and Bol, J. F. (1988): Transgenic tobacco plants accumulating tobacco rattlevirus coat protein resist infection with tobacco rattle virus and pea early browning virus.Virology 167: 649-652.
van Dun, C. M. P., Overduin, B., van Vioten-Doting, L. and Bol, J. (1988): Transgenic tobaccoexpressing tobacco streak virus or mutated alfalfa mosaic virus coat protein does not cross-protect against alfalfa mosaic virus infection. Virology 164: 383-389.
Edwardson, J. R. and Christie, R. G. (1997): Viruses infecting peppers and other solanaceouscrops. Volume II, Florida Agricultural Experiment Station, Monograph 18-II.

96
Ellis, P., Stace-Smith, R., de Viliers, G. (1997): Identification and geographic distribution ofserotypes of potato virus Y. Plant Dis. 81: 481-484.
English, J. J., Mueller, E. and Baulcombe, D. C. (1996): Supression of virus accumulation intransgenic plants exhibiting silencing of nuclear genes. Plant Cell 8: 178-188.
Falk, B. W. and Bruening, G. (1994): Will transgenic crops generate new viruses and newdiseases? Science 263: 1395-1396.
Fang, G. and Grumet, R. (1993): Genetic engineering of potyvirus resistance using constructsderived from zucchini yellow mosaic virus coat protein gene. Mol. Plant-Microbe Interact. 6:358-367.
Farinelli, L. (1992): Coat protein gene-mediated resistance to the potato virus Y /PVY/ intobacco and potato. (Thesis) 1-121.
Farinelli, L., Malnoë, P. and Collet, G. F. (1992): Heterologous encapsidation of potato virus Ystrain O (PVYO) with the transgenic coat protein of PVY strain N (PVYN) in Solanumtuberosum cv. Bintje. Bio/Technology 10: 1020-1025.
Farinelli, L. and Malnoë, P. (1993): Coat protein gene-mediated resistance to potato virus Y intobacco: examination of the resistance mechanisms -is the transgenic coat protein required forprotection? Mol. Plant-Microbe Interact. 6: 284-292.
Fehér, A., Skyrabin, K. G., Balázs, E., Preiszner, J., Shulga, O. A., Zakharyev, V. M. andDudits, D. (1992): Expression of PVX coat protein gene under the control of extensin-genepromoter confers virus resistance on transgenic potato plants. Plant Cell Rep. 11: 48-51.
Fitch, M. M. M., Manshardt, R. M., Gonsalves, D., Slightom, J. L., Sanford, J. C. (1992): Virusresistant papaya plants derived from tissues bombarded with the coat protein gene of papayaringspot virus. Bio/Technology 10: 1466-1472.
Gerber, M. and Sarkar, S. (1989): The coat protein of tobacco mosaic virus does not play asignificant role for cross-protection. J. Phytopathology 124: 323-333.
Gielen, J. J. L., De Haan, P., Kool, A. J., Peters, D., Van Grinsven, M. W. J. M. and Goldbach,R. W. (1991): Engineered resistance to tomato spotted wilt virus, a negative-strand RNA virus.Bio/Tecnology 9: 1363-1367.
Goldbach, R. and de Haan, P. (1993): Prospects of engineered forms of resistance againsttomato spotted wilt virus. Seminars in Virology 4: 381-387.
Gonsalves, D., Chee, P., Provvidenti, R., Seem, R. and Slightom, J. L. (1992): Comparison ofcoat protein-mediated and genetically-derived resistance in cucumbers to infection by cucumbermosaic virus under field conditions with natural challenge inoculations by vectors.Bio/Technology 10: 1562-1570.
Gooding, G. V. and Tolin, S. A. (1973): Strains of potato virus Y affecting flue-cured tobacco inthe southeastern United States. Plant Dis. Rep. 57:200-204.

97
Goodwin, J., Chapman, K., Swaney, S., Parks, T. D., Wernsman, E. A. and Dougherty, W. G.(1996): Genetic and biochemical dissection of transgenic RNA-mediated virus resistance. PlantCell 8: 95-105.
de Haan, P., Gielen, J. J., Prins, M., Wijkamp, I. G., van Schepen, A., Peters, D., van Grinsven,M. Q. and Goldbach, R. (1992): Characterization of RNA-mediated resistance to tomato spottedwilt virus in transgenic tobacco plants. Bio/Technology (NY) 10: 1133-1137.
Haldeman-Cahill, R., Daros, J. A. and Carrington, J. C. (1998): Secondary structures in thecapsid protein coding sequence and 3’ nontranslated region involved in amplification of thetobacco etch virus genome. Journal of Virology 72: 4072-4079.
Halk, E. L., Merlo, D. J., Liao, L.W., Jarvis, N. P. and Nelson, S. E. (1989): Resistance to alfalfamosaic virus in transgenic tobacco and alfalfa. In: Staskawicz, B., Ahlquist, P., Yolder, O.(eds.) Molecular biology of plant-pathogen interactions, Alan R. Liss, New York.
Hammond, J. and Kamo, K. K. (1993): Resistance to bean yellow mosaic virus /BYMW/ andother potyviruses in transgenic plants expressing BYMW antisense RNA, coat protein orchimera coat proteins. Acta Horticult.
Hammond, J. and Kamo, K. K. (1995): Effective resistance to potyvirus infection conferred byexpression of antisense RNS in transgenic plants. Mol. Plant-Microbe Interact. 8: 674-682.
Harrison, B. D. and Roberts, I. M. (1971): Pinwheels and crystalline structures induced byatropa mild mosaic virus with particles 925 nm long. J. gen. Virology 10: 71-78.
Hayakawa, T., Zhu, Y., Itoh, K., Kimura, Y., Izawa, T., Shimamoto, K. and Toryama, S. (1992):Genetically engineered rice resistant to rice stripe virus, an insecttransmitted virus. Proc. Natl.Acad. Sci. USA 89: 9865-9869.
Hemenway, C., Fang, R-X., Kaniewski, W. K., Chua, N-H. and Tumer, N. E. (1988): Analysis ofthe mechanism of protection in transgenic plants expressing the potato virus X coat protein orits antisense RNA. EMBO J: 7: 1273-1280.
Hill, K. K., Jarvis-Eagan, N., Halk, E. L., Krahn, K. J., Liao, L. W., Mathewson, R. S., Merlo,D. J., Nelson, S. E., Rakha, K. E. and Loesch-Fries, L. S. (1991): The development of virusresistant alfalfa, Medicago sativa L. Bio/Technology 8: 373-377.
Hoekema, A., Huisman, M. J., Molendijk, L., van den Elzen, P. J. M. and Cornelissen, B. J. C.(1988): The genetic engineering of two commercial potato cultivars for resistance to potatovirus X. Bio/Tecnology 7: 273-278.
Hollings, M. and Brunt, A. A. (1981): Potyvirus group. CMI/AAB Description of Plant Viruses#245: 7pp.
Horikoshi, M., Nakayama, M., Yamaoka, N., Furosawa, I. and Shishyama, J. (1987): Bromemosaic virus coat protein inhibits viral RNA synthesis in vitro. Virology 158: 15-19.

98
Horváth, J. (1966): Studies on strains of potato virus Y. 2. Normal strain. Acta Phytopath.Acad. Sci. Hung. 1: 333-352.
Horváth, J. (1995): A szántóföldi növények betegségei. Mezőgazda kiadó, Budapest 1996, 328pp.
Ishida, B.K., Snyder, G.W. and Belknap, W.R. (1989): The use of in vitro-grown microtuberdiscs in Agrobacterium-mediated transformation of Russet Burbank and Lehmi Russet potatoes.Plant Cell Reports 8:325-328.
Jacquet, C., Delecolle, B., Raccah, B., Lecoq, H., Dunez, J. and Ravelonandro, M. (1998a): Useof modified plum pox virus coat protein genes developed to limit heteroencapsidation-associated risks in transgenic plants. Journal of General Virology 79: 1509-1517.
Jacquet, C., Ravelonandro, M. and Dunez, J. (1998b): High resistance and control of biologicalrisks in transgenic expressing modified plum pox virus coat protein. Acta Virol. 42. (4): 235-237.
Jongedijk, E., de Schutter, A. A. J. M., Stolte, T., van den Elzen, P. J. M. and Cornelissen, B. J.C. (1992): Increased resistance to potato virus X and preservation of cultivar properties intransgenic potato under field conditions. Bio/Tecnology 10: 422-429.
Kallerhoff, J., Perez, J., Bouzoubaa, S., Bentahar, S. and Perret, J. (1990): Beet necrotic yellowvein virus coat protein-mediated protection in sugarbeet /Beta vulgaris/ protoplasts. Plant CellRep. 9: 224-228.
Kaniewski, W., Lawson, C., Sammons, B., Haley, L., Hart, J., Delannay, X. and Tumer, N. E.(1990): Field resistance of transgenic Russet Burbank potato to effects of infection by potatovirus X and potato virus Y. Bio/Technology 8: 750-754.
Kasschau, K. D., Cronin, S. and Carrington, J. C. (1997): Genome amplification and long-distance movement functions associated with the central domain of tobacco etch potyvirushelper component-proteinase. Virology 228: 251-262.
Kawchuk, L. M., Martin, R. R. and McPherson, J. (1990): Resistance in transgenic potatoexpressing the potato leafroll virus coat protein gene. Mol. Plant-Microbe Interact. 3: 301-307.
Kawchuk, L. M., Martin, R. R. and McPherson, J. (1991): Sense and antisense RNA-mediatedresistance to potato leafroll virus in Russet Burbank potato plants. Mol. Plant-Microbe Interact.4: 247-253.
Kollár, Á., Thole, V., Dalmay, T., Salamon, P. and Balázs, E. (1993): Efficient pathogen-derived resistance induced by integrated potato virus Y coat protein gene in tobacco. Biochimie75: 623-629.
Kunik, T., Salomon, R., Zamir, D., Navot, N., Zeidan, M., Michelson, I., Gafni, Y. and Chosnek,H. (1994): Transgenic tomato plants expressing the tomato yellow leaf curl virus capsid proteinare resistant to the virus. Bio/Technology 5: 500-504.

99
Laín, S., Riechmann, J. L., Méndez, E. and García, J. A. (1988): Nucleotid sequence of the 3’terminal region of plum pox virus RNA. Virus res. 10: 325-342.
Lawson, C., Kaniewski, W., Haley, L., Rozman, R., Newell, C., Sanders, P. and Turner, N.(1990): Engineering resistance to mixed virus infection in a commercial potato cultivar:resistance to potato virus X and potato virus Y in transgenic Russet Burbank. Bio/Technology8: 127-134.
Le Romancer, M., Kerlan, C., Nedellec, M. (1992): Potato necrotic ringspot disease: a geneticapproach of the phenomenon and studies about hypersensitive or extreme susceptible behaviourof several cultivars. Proceedings of the EAPR Virology Section Meeting, 91-95.
Leclerc, D. and AbouHaidar, M. G. (1995): Transgenic tobacco plants expressing a truncatedform of the PAMV capsid protein (CP) gene show CP-mediated resistance to potato aucubamosaic virus. Mol. Plant-Microbe Interact. 1: 58-65.
Lecoq, H., Ravelonandro, M., Wipf-Scheibel, C., Monsion, M., Raccah, B. and Dunez, J. (1993):Aphid transmission of a non-aphid-transmissible strain of zucchini yellow mosaic potyvirusfrom transgenic plants expressing the capsid protein of plum pox potyvirus. Mol. Plant-MicrobeInteract. 3: 403-406.
Leiser, R.-M. and Richter, J. (1978): Reinigung und einige Eigenschaffen des Kartoffel-Y-Virus. Arch. Phytopathol. u. Pflanzenschutz 14:337-350.
Li, H. W., Lucy, A. P., Guo, H. S., Li, W. X., Ji, L. H., Wong, S. M. and Ding, S. W. (1999):Strong host resistance targeted against a viral suppressor of the plant gene silencing defencemechanism. EMBO J. 18: 2683-2691.
Lindbo, J. A. and Dougherty, W. G. (1992a): Pathogen-derived resistance to a potyvirus:immune and resistance phenotypes in transgenic tobacco expressing altered forms of apotyvirus coat protein nucleotide sequence. Mol. Plant-Microbe Interact. 5: 144-153.
Lindbo, J. A. and Dougherty, W. G. (1992b): Untranslatable transcripts of the tobacco etch viruscoat protein gene sequence can interfere with tobacco etch virus replication in transgenic plantsand protoplasts. Virology 189: 725-733.
Lindbo, J. A., Silva-Rosales, L. and Dougherty, W. G. (1993a): Pathogen derived resistance topotyviruses: working, but why? Seminars in Virology 4: 369-379.
Lindbo, J. A., Silva-Rosales, L., Proebsting, W. M. and Dougherty, W. G. (1993b): Induction ofhighly specific antiviral state in transgenic plants: Implications for regulation of geneexpression and virus resistance. Plant Cell 5: 1749-1759.
Lindbo, J. A., Fitzmaurice, W. P. and della-Chioppa, G. (2001): Virus-mediated reprogrammingof gene expression in plants. Current Opin. in Plant Biology 4:181-185.
Ling, K., Namba, S., Gonsalves, C., Slightom, J. L. and Gonsalves D. (1991): Protection againstdetrimental effects of potyvirus infection in transgenic tobacco plants expressing the papayaringspot virus coat protein gene. Bio/Technology 9: 752-758.

100
Loesch-Fries, L. S., Merlo, D., Zinnen, T., Burhop, L., Hill, K., Krahn, K., Jarvis, N., Nelson, S.and Halk, E. (1987): Expression of alfalfa mosaic virus RNA 4 in transgenic plants confersvirus resistance. EMBO J. 6: 1845-1851.
Lomonossoff, G. P. (1995): Pathogen-derived resistance to plant viruses. Annual Review ofPhytopathology 33: 323-343.
López-Moya, J. J. and Pirone, T. P. (1998): Charge changes near the N-terminus of the coatprotein of two potyviruses affect virus movement. J. Gen. Virol. 79: 161-165.
MacKenzie, D. J. and Tremaine, J. H. (1990): Transgenic Nicotiana debneyi expressing viralcoat protein are resistant to potato virus S infection. J. Gen. Virology 71: 2167-2170.
MacKenzie, D. J., Tremaine, J. H. and Pherson, J. (1991): Genetically engineered resistance topotato virus S in potato cultivar Russet Burbank. Mol. Plant-Microbe Interact. 4: 95-102.
MacKenzie, D. J. and Ellis, P. J. (1992): Resistance to tomato spotted wilt virus infection intransgenic tobacco expressing the viral nucleocapsid. Mol. Plant-Microbe Interact. 5: 34-40.
Mahayan, S., Dolja, V. V. and Carrington, J. C. (1996): Roles of the sequence encoding tobaccoetch virus capsid protein in genome amplification: requirements for the translation process anda cis-active element. J. Virol. 70: 4370-4379.
Maia, I. G., Haenni, A. L. and Bernardi, F. (1996): Potyviral HC-Pro: a multifunctional portein.Journal of General Virology 77: 1335-1341.
Mäki-Valkama, T., Valkonen, J. P. T., Kreuze, J. F. and Pehu, E.( 2000): Transgeinc resistanceto PVYO associated with post-transcriptional silencing of P1 transgene is overcome by PVYN
strains that carry highly homologous P1 sequences and recover transgene expression atinfection. Mol. Plant-Microbe Interact. 4:366-373.
Mallory, A. C. (2001): HC-Pro suppression of gene silencing eliminates the small RNAs but notthe transgene methylation or the mobile signal. Plant Cell 13: 571-583.
Malnoë, P., Farinelli, W., Collet, G. F. and Reust, W. (1994): Small-scale field tests withtransgenic potato, cv. Bintje, to test resistance to primary and secondary infections with potatovirus Y. Plant Mol. Biol. 25: 936-975.
Marathe, R., Anandalakshmi, R., Smith, T. H., Pruss, G. J. and Vance, V. (2000): RNA virusesas inducers, suppressors and targets of post-transciptional gene silencing. Plant Mol. Biol. 43:295-306.
Matzke, M. A., Matzke, A. J. M., Pruss G. J. and Vance V. B. (2001): RNA-based silencingstrategies in plants. Curr. Opin. in Genetics and Development 11:211-227.
McKinney, H. H. (1929): Mosaic disease in the Canary Islands, West Africa and Gibraltar.Journal of Agricultural Researche 39: 557-558.

101
Mueller, E., Gilbert, J. E., Davenport, G., Brigneti, G. and Baulcombe, D. C. (1995):Homology-dependent resistance: Transgenic virus resistance in plants related to homology-dependent gene silencing. Plant Journal 7: 1001-1013.
Murashige, T. and Skoog, F. (1962): A revised medium for rapid growth and bioassay withtobacco tissue culture. Physiol. Plant. 15: 473-497.
Murphy, J. F., Hunt, A. G., Rhoads, R. E. and Shaw, J. G. (1990): Expression of potyviral genesin transgenic tobacco plants. Abstr. VIII. Int. Congr. Virol. Berlin, aug. 26-31. P84-007.
Murphy, J. F., Rhoads, R. E., Hunt, A. G.and Shaw, J. G. (1990): The VPg of tobacco etch virusRNA is the 49- kDa proteinase or the N-terminal 24 kDa part of the proteinase. Virology 178:285-288.
Murry, L. E., Elliot, L. G., Capitant, S. A., West, J. A., Hanson, K. K., Scarafia, L., Johnston, S.,DeLuca-Flathery, C., Nichols, S., Cuanan, D., Dietrich, P. S., Mettler, S. D., Warnick, D. A.,Rhodes, C., Shinibaldi, R. M. and Brunke, K. J. (1993): Transgenic corn plants expressingMDMV strain B coat protein are resistant to mixed infections of maize dwarf mosaic virus andmaize chlorotic mottle virus. Bio/Technology 11: 1559-1564.
Nakajima, M., Hayakawa, T., Nakamura, I. and Suzuki, M. (1993): Protection against cucumbermosaic virus /CMV/ strains O and Y and chrisanthemum mild mottle virus in transgenictobacco plants expressing CMV-O coat protein. J. Gen. Virol. 74: 319-322.
Namba, S., Ling, K., Gonsalves, C., Gonsalves, D. and Slightom, J. L. (1991): Expression of thegene encoding the coat protein of cucumber mosaic virus /CMV/ strain WL appears to provideprotection to tobacco plants against infection by several different CMV strains. Gene 107: 181-188.
Namba, S., Ling, K., Gonsalves, C., Gonsalves, D. and Slightom, J. L. (1992): Protection oftransgenic plants expressing the coat protein gene of watermelon mosaic virus II or zucchiniyellow mosaic virus against six potyviruses. Phytopathology 82: 940-946.
Nejidat, A. and Beachy, R. N. (1990): Transgenic tobacco plants expressing a coat protein geneof tobacco mosaic virus are resistant to some other tobamoviruses. Mol. Plant-Microbe Interact.3: 247-251.
Nelson, R. S., McCormick, S. M., Delannay, X., Dubé, P., Layton, J., Anderson, E. J., Kaiewski,M., Proksch, R. K., Horsch, R. B., Rogers, S. G., Fraley, R. T. and Beachy, R. N. (1988): Virustolerance, plant growth and field performance of transgenic tomato expressing coat protein fromtobacco mosaic virus. Bio/Technology 6: 403-409.
Okuno, T., Nakayama, M., Yoshida, S., Furusawa, I. and Koyima, T. (1993): Comparativesusceptibility of transgenic tobacco plants expressing the coat protein gene of cucumber mosaicvirus to infection with virions and RNA. Phytopathology 83: 542-547.
Palauqui, J. C., Elmayan, T., Pollien, J. M. and Vaucheret, H. (1997): Systemic acquiredsilencing: transgene-specific post-transcriptional silencing is transmitted by grafting fromsilenced stocks to non-silenced scions. EMBO J. 15: 4738-4745.

102
Palauqui, J. C. and Balzergue, S. (1999): Activation of systemic acquired silencing by localisedintroduction of DNA. Current Biology 9: 59-66.
Palkovics, L., Wittner, A. and Balázs, E. (1995): Pathogen-derived resistance induced byintegrating the plum pox virus coat protein gene into plants of Nicotiana benthamiana. ActaHorticulturae 386:311-317.
Palkovics, L., Karamova, N., Pribék, D. and Balázs. E. (1998): Changes in the 5’ non-codingregion of plum pox virus in connection with symptom development. 7th International Congressof Plant Pathology, Edinburgh, Scotland; Abstracts, Vol. 2./ 1.11.34.
Palukaitis, P. and Zaitlin, M. (1984): A model to explain the "crossprotection" phenomenonshown by plant viruses and viroids, in "Plant-Microbe Interactions. Molecular and GeneticPerspectives." Vol.1. (T. Kosuge and E. W. Nester, eds.) Macmillan publishing company, NewYork. pp. 420-429.
Pang, S. Z., Slightom, J. L. and Gonsalves, D. (1993): Different mechanisms protect transgenictobacco against tomato spotted wilt and impatiens necrotic spot tospoviruses. Bio/Technology11: 819-824.
Pirone, T. P. (1991): Viral genes and gene products that determine insect transmissibility.Semin. Virol. 2: 81-87.
Pirone, T. P. and Blanc, S. (1996): Helper-dependent vector transmission of plant viruses.Annu. Rev. Phytopathol. 34: 227-247.
Powell-Abel, P., Nelson, R. S., De, B., Hoffmann, N., Rogers, S. G., Fraley, R. T. and Beachy, R.N. (1986): Delay of disease development in transgenic plants that express the tobacco mosaicvirus coat protein gene. Science 232: 738-743.
Powell-Abel, P., Sanders, P. R., Turner, N., Fraley, R. T. and Beachy, R. N. (1990): Protectionagainst tobacco mosaic virus infection in transgenic tobacco plants requires accumulation ofcoat protein rather than coat protein RNA sequences. Virology 175: 124-130.
Prins, M., de Haan, P., Luyten, R., van Heller, M., van Grinsven, M. Q. J. M. and Goldbach, R.(1995): Broad resistance to tospoviruses in transgenic tobacco plants expressing three tospoviralnucleoprotein sequences. Mol. Plant-Microbe Interact. 1: 85-91.
Pruss, G., Ge, X., Shi, X. M., Carrington, J. C. and Bowman Vance, V. (1997): Plant viralsynergism: the potyviral genome encodes a broad-range pathogenicity enhancer thattransactivates replication of heterologous viruses. The Plant Cell 9: 859-868.
Quemada, H. D., Gonsalves, D. and Slightom, J. L. (1991): Expression of coat protein genefrom cucumber mosaic virus strain C in tobacco: Protection against infection by CMV strainstransmitted mechanically or aphids. Phytopathology 81: 794-802.
Ratcliff, F., Bryan, D. H. and Baulcombe, D. C. (1997): A similarity between viral defense andgene silencing in plants. Science 276: 1558-1560.

103
Ravelonandro, M., Monsion, M., Delbos, R. and Dunez, J. (1993): Variable resistance to plumpox virus and potato virus Y infection in transgenic Nicotiana plants expressing plum pox viruscoat protein. Plant Science 91: 157-169.
Regner, F., da Camara Machado, A., da Camara Machado, M. L., Steinkeller, H., Mattanovich,D., Hanzer, H., Weiss, H. and Katinger, H. (1992): Coat protein mediated resistance to plumpox virus in Nicotiana clevelandii and N. benthamiana. Plant Cell Rep. 11: 30-33.
Reimann-Philip, U. and Beachy, R. N. (1993): The mechanism(s) of coat protein-mediatedresistance against tobacco mosaic virus. Seminars in Virology 6: 349-357.
Riechmann, J. L., Laín, S. and García, J. A. (1990): Infectious in vitro transcripts from a plumpox potyvirus cDNA clone. Virology, 177: 710-716.
Riechmann, J. L., Lain, S. and Garcia, J. A. (1992): Highlights and prospects of potyvirusmolecular biology. J. Gen. Virology 73: 1-16.
Riechmann, J. L., Cervera, M. T. and Garcia, J. A. (1995): Processing of the plum pox viruspolyprotein at the P3-6K1 junction is not required for virus viability. Journal of GeneralVirology 76: 951-956.
Robaglia, C., Durand-Tardif, M., Trochet, M., Boudazin, G., Astier-Manifavier, S. and Casse-Delbart, F. (1989): Nucleotide sequence of potato virus Y (N strain) genomic RNA. J. Gen.Virol. 70: 935-947.
Rodriguez-Cerezo, E., Ammar, E. D., Pirone, T. P. and Shaw, J. G. (1993): Association of thenon-structural P3 viral protein with cylindrical inclusions in potyvirus-infected cells. Journal ofGeneral Virology 74: 587-592.
Rodriguez-Cerezo, E., Findlay, K., Shaw, J. G., Lomonossoff, G. P., Qiu, S. G., Linstead, P.,Shanks, M. and Risco, C. (1997): The coat and cylindrical inclusion proteins of a potyvirus areassociated with connections between plant cells. Virology 236: 296-306.
Rojas, M. R., Zerbini, F. M., Allison, R. F., Gilbertson, R. L. and Lucas, W. J. (1997): Capsidprotein and helper component-proteinase function as potyvirus cell-to-cell movement proteins.Virology 237:283-295.
Rubino, L., Carrington, J. C., Russo, M. (1992): Biologically active cymbidium ringspot virussatellite RNA in transgenic plants suppresses accumulation of DI RNA. Virology 188: 429-437.
Rubino, L., Caprioti, G., Lupo, R. and Russo, M. (1993): Resistance to cymbidium ringspottombusvirus infection in transgenic Nicotiana benthamiana plants expressing the virus coatprotein gene. Plant Mol. Biol. 21: 665-672.
Salomon, R., Raccah, B. (1990): The role of the N-terminus of potyvirus coat protein in aphidtransmission. VIII. International Congress of Virology, Berlin. Abstract p83-87.

104
Salomon, R. and Bernardi, F. (1995): Inhibition of viral aphid transmission by the N-terminusof the maize dwarf mosaic virus coat protein. Virology 213: 676-679.
Sambrook, J., Fritsch, E.F. and Maniatis, T. (1989): Molecular cloning: Laboratory Manual, 2nd
ed. (Cold Spring Harbor, NY: Cold Spring Harbor Laboratory)
Sanders, P. R., Sammons, B., Kaniewski, W., Haley, L., Layton, J., Lavallee, B. J., Delannay, X.and Tumer, N. E. (1992): Field resistance of transgenic tomatoes expressing the tobacco mosaicvirus or tomato mosaic virus coat protein genes. Phytopathology 82: 683-690.
Sanford, J. C. and Johnston, S. A. (1985): The concept of pathogen derived resistance: Derivingresistance genes from the parasites own genome. J. Theor. Biol. 113: 395-405.
Sarkar, S., Smitamana, P. (1981): A proteinless mutant of tobacco mosaic virus: evidenceagainst the role of a viral coat protein for interference. Mol. Gen. Genet. 184: 158-159.
Scorza, R., Cordts, J. M., Mante, S., Gonsalves, D., Damsteegt, V. D., Yepes, L. M. andSlightom, J. L. (1991): Agrobacterium-mediated transformation of plum (Prunus domestica L.)with the coat protein gene of papaya ringspot virus. Third Int. Congr. Plant Mol. Biol. 6-11.Oct. 1991. Tuscon, AZ. abstract 1167.
Scorza, R., Ravelonandro, M., Callahan, A. M., Cordts, J. M., Fuchs, M., Dunez, J. andGonsalves, D. (1994): Transgenic plums (Prunus domestica L.) express the plum pox virus coatprotein gene. Plant Cell Reports 14: 18-22.
Sherwood, J. L., Fulton, R. W. (1982): The specific involvement of coat protein in tobaccomosaic virus cross protection. Virology 119: 150-158.
Shukla, D. D., and Ward, C. W. (1989): Structure of potyvirus coat proteins and its applicationin the taxonomy of the potyvirus group. Adv. Virus Res. 36: 273-314.
Shukla, D. D., Frenkel, M. J. and Ward, C W. (1991): Structure and function of the potyvirusgenome with special reference to the coat protein coding region. Can. J. Plant Pathology 13:178-191.
Shukla, D. D., Ward, C. W., Brunt, A. A. (1994): The Potyviridae. CABI, University Press,Cambridge UK
Sijen, T. and Kooter, J. M. (2000): Post-transcritional gene-silencing: RNAs on the attack or onthe defense? Bioessays 22: 520-531.
Smith, K. M. (1931): Composite nature of certain potato viruses of the necrotic group. Nature127, 702.
Smith, K. M. and Dennis, R. W. G. (1940): Some notes on a suspected variant of Solanum virus2 (potato virus Y). Ann. Appl. Biol. 27: 65-70.

105
Smith, H. A., Swaney, S. L., Parks, T. D., Wernsam, E. A. and Dougherty, W. G. (1994):Transgenic plant virus resistance mediated by untranslatable sense RNAs: expression,regulation, and fate of nonessential RNAs. Plant Cell 6: 1441-1453.
Smith, H. A., Powers, H., Swaney, S. L., Brown, C. and Dougherty (1995): Transgenic potatovirus Y resistance in potato: Evidence for an RNA-mediated cellular response. Phytopathology85: 864-870.
Sonoda, S., Mori, M. and Nishiguchi, M. (1999): Homology-dependent virus resistance intransgenic plants with the coat protein gene of sweet potato feathery mottle potyvirus: Targetspecificity and transgene methylation. Virology 5:385-391.
Soumounon, Y. and Laliberté, J. F. (1994): Nucleic acid-binding properties of the P1 protein ofturnip mosaic potyvirus produced in Echerichia coli. Journal of General Virology 75:2567-2573.
Stark, D. M. and Beachy, R. N. (1989): Protection against potyvirus infection in transgenicplants: evidence for a broad spectrum resistance. Bio/Technology 7: 1257-1262.
Sudarsono- Woloshuk, S. L., Xiong, Z., Hellmann, G. M., Wernsman, E. A., Weissinger, A. K.and Lommel, S. A. (1993): Nucleotide sequence of the capsid protein cristons from six potatovirus Y (PVY) isolates infecting tobacco. Arch. Virol. 132:161-170.
Szirmai, J. (1958): A burgonya Y-vírusának érbarnulást okozó változata a dohánykultúrákban.Növénytermelés 7: 341-350.
Tacke, E., Salamini, F. and Rohde, W. (1996): Genetic engineering of potato for broad-spectrumprotection against virus infection. Nat. Biotechnology 14: 1579-1601.
Takács, A. P. (2001): A burgonya Y vírus (potato Y potyvirus, PVY) jellemzése, az NTNtörzzsel szembeni rezisztencia a vad Solanum fajokban. Doktori (PhD) Értekezés
Tepfer, M. (1993): Viral genes and transgenic plants (What are the potential environmentalrisks?) Bio/Technology 11: 1125-1129.
Tepfer, M. and Balázs, E. (1997): Virus-resistant Transgenic Plants: Potential EcologicalImpact. 1997. Springer-Verlag, Berlin, Heidelberg, INRA Paris.
Thole, V., Dalmay, T., Burgyán, J. and Balázs, E. (1993): Cloning and sequencing of potatovirus Y (Hungarian isolate) genomic RNA. Gene
Thole, V. (1993): A magyarországi flórából származó burgonya Y vírus teljes genomjánakmolekuláris klónozása, nukleotid sorrendjének meghatározása és mesterséges vírus rezisztenciakialakítása. Kandidátusi értekezés. 1-103.
Tumer, N. E., O'Conell, K. M., Nelson, R. S., Sanders, P. R., Beachy, R. N., Fraley, R. T. andShah, D. M. (1987): Expression of alfalfa mosaic virus coat protein gene confers cross-protection in transgenic tobacco and tomato plants. EMBO J. 6: 1181-1188.

106
Tumer, N. E., Kaniewski, W., Haley, L., Gehrke, L., Lodge, J. K. and Sanders, P. (1991): Thesecond amino acid of alfalfa mosaic virus coat protein is critical for coat protein-mediatedprotection. Proc. Natl. Acad. Sci. USA 88: 2331-2335.
Vaira, A. M., Semeria, L., Crespi, S., Lisa, V., Allavena, A. and Accotto, G. P. (1995):Resistance to tospoviruses in Nicotiana benthamiana transformed with the N gene of tomatospotted wilt virus: Correlation between transgene expression and protection in primarytransformants. Mol. Plant-Microbe Interact. 1: 66-73.
Van den Heuvel, J. F. J. M., van der Vlugt, R. A. A., Verbeck, M. N., den Haag, P. T. andHuttinga, H. (1994): Characteristics of a resistance-breaking strain of potato virus Y causingpotato tuber necrotic ringspot disease. European Journal os Plant Pathology, 100: 347-356.
Varrelmann, M. and Maiss, E. (2000): Mutations in the coat protein gene of plum pox virussuppress particle assembly, heterologous encapsidation and complementation in transgenicplants of Nicotiana benathamiana. J. Gen. Virol. 81: 567-576.
Vaucheret, H., Beclin, C., Elmayan, T., Feuerbach, F., Godon, C., Morel, J. B., Mourrain, P.,Palauqui, J. C. and Vernhettes, S. (1998): Transgene-induced gene silencing in plants. PlantJournal 16: 651-659.
Vaucheret, H. and Vance, V. (2001): RNA silencing in plants- defence and counterdefence.Science, 292: 2277-2280.
Verchot, J., Koonin, E. V. and Carrington, J. C. (1991): The 35-kDa protein from the N-terminus of the potyviral polyprotein functions as a third virus-encoded proteinase. Virology185: 527-535.
van der Vlugt, R. A. A., Ruiter, K. R. and Goldbach, R. (1992): Evidence for sense RNA-mediated protection to PVYN in tobacco plants transformed with the viral coat protein criston.Plant Mol. Biol. 20: 631-639.
Voinnet, O. (2001): RNA silencing as a plant immune system against viruses. Trends inGenetics, 17, 8:449-459.
Voinnet, O., Vain, P., Angell, S. and Baulcombe, D. (1998): Systemic spread of sequence-specific transgene RNA degradation in plants is inhibited by localised introduction of ectopicpromoterless DNA. Cell 95: 177-187.
Voinnet, O., Pinto, Y. M. and Baulcombe, D. C. (1999): Suppression of gene silencing: a generalstrategy used by diverse DNA and RNA viruses of plants. Proc. Natl. Acad. Sci. USA 96:14147-14152.
Ward, C. W. and Shukla, D. D. (1991): Taxonomy of potyviruses: Current problems and somesolutions. Intervirology 32: 269-296.
Waterhouse, P. M., Wang, M-B and Finnegan, E. J. (2001a): Role of short RNAs in genesilencing. Trends in Plant Science 6; 7: 297-301.

107
Waterhouse, P. M., Wang, M-B. and Lough, T. (2001b): Gene silencing as an adaptive defenseagainst viruses. Nature, 411:834-842.
White, J. L. and Kaper, J. M. (1989): A simple method for detection of viral satellite RNAs insmall plant tissue sampls. J. Virol. Methods 23: 83-94.
van der Wilk, F. P.-L., Willink, D., Huisman, M. J., Huttinga, H. and Goldbach, R. (1991):Expression of potato leafroll luteovirus coat protein gene in transgenic potato plants inhibitsviral infection. Plant Mol. Biol. 17: 431-439.
Wilson, T. M. A. (1993): Strategies to protect crop plants against viruses: Pathogen-derivedresistance blossoms. Proceedings of the National Academy of Sciences USA 90: 3134-3141.
Wisniewski, L. A., Powell, P. A., Nelson, R. S., Beachy, R. N. (1990): Local and systemic spreadof tobacco mosaic virus in transgenic tobacco plants. Plant Cell 2: 559-567.
Wolf, I. és Horváth, S. (2000): A burgonya Y vírus (potato Y potyvirus, PVY) törzseinekelőfordulása burgonya-termőterületeken Magyarországon. Növényvédelem 36, 449-453.
Xu, X. and Shaw, J. G. (1998): Evidence that assembly of a potyvirus begins near the 5’terminus of the viral RNA. Journal of General Virology 79: 1525-1529.
Yushibov, V. and Loesch-Fires, L. S. (1995): High-affinity RNA-binding domains of alfalfamosaic virus coat protein are not required for coat protein-mediated resistance. Proc. Natl.Acad. Sci. USA 92: 8980-8984.
Zaitlin, M. (1976): Viral cross protection: more understanding is needed.Phytopathology 66: 382-383.
Zambryski, P., Joos, H., Gentello, C., Leemans, J., Van Montagu, M. and Shell, J. (1983): Ti-plasmid vector for the introduction of DNA into cells without altering their normal regenerationcapacity. The EMBO Journal 2:2143-2150.
Zinnen, T. M., Fulton, R. W. (1986): Cross-protection between sunn-hemp mosaic and tobaccomosaic viruses. J. Gen. Virology 67: 1679-1697.
de Zoeten, G. A., Fulton, R. W. (1975): Understanding generates possibilities. Phytopathology65: 221-222.

108
A disszertációhoz kapcsolódó tudományos közlemények
jegyzéke:
Józsa, R., Stasevski, Z., Wolf, I., Horváth, S. and Balázs, E. (2002): Potatovirus Y coat protein gene induced resistance in valuable potato cultivars. ActaPhytopathologica et Entomologica 1-2.
Józsa, R., Stasevski, Z. and Balázs, E. (2002): High level of field resistance oftransgenic tobaccoes induced by integrated potato virus Y coat protein gene.Acta Phytopathologica et Entomologica Hungarica 37 311-316.
Józsa, R., Balázs, E. (2000): Víruseredetű köpenyfehérje gének által közvetítettrezisztencia transzgénikus növényekben I. (Helyzetkép az ezredfordulón)Review, Növénytermelés 49, No. 1-2: 165-184.
Józsa, R., Balázs, E. (2000): Víruseredtű köpenyfehérje gének által közvetítettrezisztencia transzgénikus növényekben II. (Hatásmechanizmusok, lehetségeskockázati tényezők) Növénytermelés 49, No. 4: 447-453.
Józsa, R. és Balázs, E. (2000): Burgonya Y vírus ellenállóság kialakításagénsebészeti úton dohány- és burgonyanövényekben. Az MTA általánosMikorbiológiai Bizottságának Tanácsadó Testületének űlése. 2000 október 4.Budapest.
Józsa, R. és Balázs, E.: Burgonya Y vírus köpenyfehérje gén beépítéseburgonyába. Növényvédelmi Tudományos Napok, Budapest 1997. február 24-25.
Szilassy, D., Józsa, R., Palkovics, L., Salánki, K., Wittner, A., Szász, A. andBalázs, E. (1997): Coat protein mediated cross protection in different plantvirus interactions. VIII th International Congress of Biotechnology, Budapest,Hungary; August 17-21. (Poster)
Horváth, S., Wolf, I., Józsa, R. és Balázs, E.: Burgonya Y-vírus köpenyfehérjegénnel transzformált burgonyavonalak rezisztenciája a burgonya Y (potato Y,PVY) potyvírussal szemben. VIII. Növénynemesítési Tudományos Napok.2002. febr. 12-13. Budapest.

109
I. Wolf, R. Józsa, E. Balázs, S. Horváth: Resistance of an old Hungarian varietytransformed with PVY-CP gene against PVY strains. European Association forPotato Research, Triennial Conference, Hamburg, july 15-19. 2002.
L. Palkovics, R. Kryldakov, J. Nádudvari, R. Józsa and E. Balázs (2003):Sequence variants of potato virus Y in field grown tobacco with differentresistance background including transgenic ones. Acta Phytopathologica etEntomologica Hungarica ( közlésre benyújtva)

110
Köszönetnyilvánítás
Köszönetet szeretnék mondani Dr. Balázs Ervinnek, a Környezet
Biotechnológiai Intézet igazgatójának, témavezetőmnek illetve Dr. Hornok
Lászlónak, programvezetőmnek, hogy megteremtették a körülményeket
kutatásaim elvégzéséhez és lehetővé tették disszertációm elkészítését.
Köszönettel tartozom kollégáimnak Dr. Wittner Anitának, Dr. Szilassy
Dénesnek, Dr. Palkovics Lászlónak és még sokan másoknak munkám során
nyújtott értékes elméleti és gyakorlati segítségükért. Köszönöm Dr. Horváth
Sándornak, Dr. Wolf Istvánnak és Dr. Nagy Gyulának, hogy lehetővé tették a
transzgénikus vonalak szántóföldi vizsgálatát. A technikai téren nyújtott
segítségükért pedig köszönet illeti Nagy Ágnest, Nádudvariné Novák Julikát és
Takács Gábort.

111
1. Melléklet
Vírusrezisztencia kiváltására irányuló kísérletek a transzgén CP eredete szerint,
vírusnemzetségenként csoportosítva
Alfamovirus nemzetség
Tumer és munkatársai (1987) a lucerna mozaik vírus (alfalfa mosaicalfamovirus –AMV) 4-es RNS-ét használták fel transzgén konstrukciókelőállításához. Dohány és paradicsom növényeket használtak atranszformációkhoz, melyek leveleiből a kivonható összfehérjékből 0.1-0.8 %-ban immunoblot analízissel mutatták ki a CP-t. A transzgénikusdohánynövényeket inokulálták 5 és 50 µg/ml koncentrációjú AMV-vel. Kéthéttel a fertőzés után a CP-t expresszáló (CP+) növényeken nem jelentek meg avírustünetek, míg a CP-t nem termelő (CP-) növényeken léziók és szisztemikusmozaik tünetek fejlődtek ki. Ezekben a fogékony növényekben 400-szornagyobb vírusmennyiséget találtak, mint a „CP+” növényekben.
Loesch-Fries és munkatársai (1987) dohány és lucerna növényekettranszformáltak kétféle génkonstrukcióval. Az egyik a karfiol mozaik vírus(cauliflower mosaic caulimovirus –CaMV) 19S promóterét és az AMV CPkonstrukciót tartalmazta, míg a másik a CaMV 35S promótert és a CP:T-DNSORF 25 3' vég szekvenciákat. A P19S:CP konstrukciót hordozódohánynövények (33 vonal) leveleiből 0.05 %-ban mutatták ki a levélösszfehérjéihez viszonyítva a CP-t, a P35S:CP konstrukcióval rendelkezőkből(34 vonal) pedig 0.13 %-ban. A P19S:CP transzgénikus utódok, melyek alegmagasabb szinten akkumulálták a CP-t, kevesebb elsődleges fertőzési tünetetmutattak, mint a „CP-” növények és ezekben a „CP+” növényekben aszisztemikus tünetek is lassabban fejlődtek ki. A 0.006 %-ban CP-t tartalmazónövények már nem voltak rezisztensek. Az ellenállóképesség hatásosnakbizonyult két AMV törzzsel szemben, de AMV RNS-el és TMV-vel történőfertőzéssel szemben már nem.
Anderson és mtsai (1989) megfigyelték, hogy az AMV CP-t tartalmazótranszgénikus növények TMV-vel szemben érzékenyek, azonban 0.01-0.5µg/ml burgonya X vírus (potato X potexvirus –PVX) vagy uborka mozaik vírus(cucumber mosaic cucumovirus –CMV) fertőzés esetén a szimptómákkifejlődésének késését tapasztalták.

112
Van Dun és mtsai (1987) lucernát transzformáltak P35S:AMV CPkonstrukcióval. A vizsgált 240 növény 40 %-a expresszálta a CP gént, mely0.05 %-ban volt jelen a levél összfehérjéiben. A transzgénikus növényekellenállóak voltak AMV fertőzéssel szemben. 5 hónappal az inokulációtkövetően sem fejlődtek ki a vírustünetek és az ezekből előállított protoplasztokis rezisztensnek mutatkoztak. Később dohánynövényeket is transzformáltakP35S:AMV CP:NOS 3' konstrukcióval. A 15 regeneráns vonalból 11-bentapasztaltak CP expressziót, melyek megközelítőleg 0.05 %-ban tartalmazták aCP-t a levél kivonható fehérjéiben. A növények rezisztensek voltak AMV'YSMV' törzzsel történő fertőzést követően, míg fogékonynak bizonyultakAMV RNS és dohány csíkosság vírus (tobacco streak ilarvirus –TSV) fertőzésesetében is. Egy "frame-shift" mutációt tartalmazó konstrukcióval transzformáltnövények (melyek csak a CP mRNS-t akkumulálták, magát a CP-t nem)fogékonyak voltak AMV fertőzéssel szemben. Ezek az eredmények afehérjetermék szerepére utalnak a rezisztencia kialakításában.
Yushibov és Loesch-Fries (1995) szintén AMV CP-vel transzformáltakNicotiana tabacum L. cv. Xanthi dohánynövényeket, melyek hatékony védelmetnyújtottak a donorvírussal szemben. Mutáns, nem-transzlálódó CP génttartalmazó konstrukciók használatát követően gyenge rezisztenciát tapasztaltak.Ez az eredmény arra mutatott rá, hogy az ellenállóképesség kialakításához nincsszükség a CP-k és az AMV RNS-ek közötti interakcióra.
Carlavirus nemzetség
Köpenyfehérje által kiváltott rezisztenciát hoztak létre a burgonya M vírus(potato M carlavirus –PVM) és burgonya S vírus (potato S carlavirus –PVS)köpenyfehérje gének felhasználásával is. MacKenzie és Tremaine (1990) ill.MacKenzie és mtsai (1991) dohánynövényekbe építették be a PVS 'An'izolátum CP génjét, mely által sikerült rezisztenciát kiváltani a transzgénikusvonalakban. Russet Burbank burgonyanövényeket transzformáltak akonstrukcióval és azt tapasztalták, hogy a CP-t expresszáló vonalak ellenállóakvoltak PVS 'An'-nel szemben, míg PVM-mel szemben kevésbé.
Cucumovirus nemzetség
Quemada és mtsai (1991) CMV 'C' törzsének CP-jével transzgénikus N.tabacum L. cv. Xanthi vonalakat állítottak elő. A rezisztencia mértéke változóvolt és függött a fertőzéshez használt törzstől (CMV 'C' subgroup 1, 'Chi' sg. 1,'WL' sg. 2). A fertőzéseket mechanikailag és levéltetvek segítségével iselvégezték. Megfigyelték, hogy a köpenyfehérjét expresszáló vonalak CMV 'C'-

113
vel szemben magas fokú rezisztenciát mutattak, míg CMV 'Chi'-vel szembenmegközelítőleg 20 %-uk illetve CMV 'WL'-lel szemben mintegy 50 %-ukmegfertőződött. A köpenyfehérjét nem termelő vonalak CMV 'C' esetén 50 %-ban fertőződtek, míg a CMV 'Chi' és 'WL' törzsek esetében 100 %-os volt afertőzöttség.
Okuno és mtsai (1993) CMV köpenyfehérje génnel transzgénikusdohánynövényeket fertőztek CMV virionnal és RNS-el. A növények mindkétesetben ellenállóak voltak, míg protoplasztok RNS-el történő fertőzésekor nemmaradt fenn az ellenállóképesség. 34 0C-on végzett hőkezeléses kísérletek soránazt tapasztalták, hogy a növényekben csökkent a rezisztencia szintje vagy akárteljesen el is tűnt. Mivel a CP magas hőmérsékleten instabil, ez az eredmény aköpenyfehérje szerepére utal CMV-vel szembeni rezisztencia kialakulásában.CMV 'D', 'WL' és 'O' izolátumok köpenyfehérje génjeivel transzformáltakdohánynövényeket Cuozzo és mtsai (1988), Namba és mtsai (1991) ésNakajima és mtsai (1993). CMV 'D' esetében védettséget tapasztaltak CMV 'C'-vel szemben, CMV 'WL' eredetű transzgént hordozóknál CMV 'C', 'Chi' és'WL'-el szemben, valamint CMV 'O' CP használatakor CMV 'O' fertőzéstkövetően.
Enamovirus nemzetség
A borsó enációs mozaik vírus (pea enation mosaic enamovirus –PEMV)CP génjét tartalmazó konstrukció felhasználásával Chowrira és mtsai (1998)vírusellenállóságot hoztak létre borsóban. Axilláris merisztémasejtekettranszformáltak injektálással és elektroporálással. A transzgénikus növényekbena kontroll növényekhez képest késői és alacsonyabb szintű PEMV replikációt,illetve gyengült tünetek kifejlődését tapasztalták.
Furovirus nemzetség
Kallerhoff és mtsai (1990) a répa nekrotikus sárgaerűség vírus (beetnecrotic yellow vein furovirus /újabban benyvirus/ -BNYVV) CP génjéthasználták fel cukorrépa protoplasztok Agrobacterium-os transzformációjára. Asejtszuszpenzióból CP-t expresszáló transzformált protoplasztokat izoláltak,melyeket fertőzve rezisztenciát tapasztaltak. Megközelítőleg a sejtek 2 %-a volttranszformáns, melyek 200 mg/l kanamicint tartalmazó szelektív táptalajon isképesek voltak növekedni. A rezisztencia magas inokulum koncentráció mellettis megmaradt és RNS-el történő fertőzés esetében sem tapasztaltakvírusreplikációt.

114
Geminivirus nemzetség
Kunik és mtsai (1994) transzgénikus paradicsomnövényeket állítottak elő aparadicsom sárga levélgöndörödés vírus (tomato yellow leaf curl geminivirus –TYLCV), amely egyszálú DNS-t tartalmazó vírus, köpenyfehérje génjénekfelhasználásával. A növények ellenállóak voltak a későbbi vírustámadássalszemben. A kísérlethez a Lycopersicon esculentum x L. pennellii hibrid F1nemzedékét használták, mivel ez a természetes paradicsommal azonosfogékonyságú, viszont a regenerációs kapacitása annál nagyobb. A CP génnel(V1) transzformált növényeket, fehérlegyek segítségével fertőzték meg. Atünetek kifejlődésének késését és recovery jelenséget is tapasztaltak. Ismételtfertőzések esetében sem mutatták ki a rezisztencia csökkenését, inkább annakerősödését figyelték meg. Kilenc transzgénikus vonalból hatban fordult előfehérje akkumuláció. A kontrollokhoz hasonlóan érzékeny transzformáltnövények csak a V1 RNS-t expresszálták, míg azok, melyek magát a CP-t istermelték, képesek voltak felépülni, vagy ellenállóságot kialakítani.
Ilarvirus nemzetség
Van Dun és mtsai (1988) TSV 'WC' törzsének CP génjével transzformáltakN. tabacum cv. Samsun és Xanthi dohánynövényeket. A növények fertőzését 10µg/ml koncentrációjú TSV inokulummal végezték. A szisztemikus szimptómáka kontroll és a „CP-” növényeken 3 héten belül kifejlődtek, míg a „CP+”vonalakban 3 hét után sem alakultak ki.
Luteovirus nemzetség
Kawchuk és mtsai (1990) burgonya levélsodródás vírus (potato leafrollluteovirus –PLRV) köpenyfehérje gént használtak Desiree, majd RussetBurbank burgonyafajták transzformációjához. A gént hordozó vonalakrezisztensnek bizonyultak PLRV fertőzéssel szemben, bár a magastranszkriptumszint ellenére alacsony köpenyfehérje mennyiséget mutattak ki:0.01 %-ot az összes levélproteinhez viszonyítva. Ennek magyarázata lehet a CPmRNS alacsony transzlációs hatékonysága vagy a protein alegységinstabilitása, de előfordulhat, hogy a köpenyfehérje csak egy bizonyosszövettípuson belül stabil, pl a floem sejtekben. A transzgénikus vonalakellenállóképességét, sem a levéltetűvel történő fertőzés, sem a magasinokulumkoncentráció nem törte át.

115
Nepovirus nemzetség
Az arabis mozaik vírus (arabis mosaic nepovirus –ArMV) és a szőlő krómmozaik vírus (grapevine chrome mosaic nepovirus –GCMV) köpenyfehérjegénjeit használták fel Brault és mtsai (1993) köpenyfehérje által indukáltkeresztvédettség kialakításához dohánynövényekben. A GCMV CP-t hordozónövények magas szinten expresszálták a köpenyfehérje gént, ami rezisztenciátis eredményezett. Néhány növény megfertőződött ugyan, de a kontrollokhozképest alacsony szinten akkumulálták a vírus RNS-t. Az ellenállóság tisztítottvírus RNS-sel történő fertőzés esetében is megmaradt. Azt tapasztalták, hogycsak a magas szintű CP expresszió eredményez csak rezisztenciát. Néhánynövény 1 µg/ml inokulum koncentrációnál is megfertőződött, míg magaskoncentrációnál (10 µg/ml) már a nagy részük. Ezek azonban jóval kevesebbRNS-t halmoztak fel, mint a szintén megfertőződött kontrollnövények. RNS-seltörténő fertőzésnél is hasonló eredményeket kaptak.
Potexvirus nemzetség
Leclerc és AbouHaidar (1995) burgonya aucuba mozaik vírus (potatoaucuba mosaic potexvirus –PAMV) köpenyfehérje génnel transzformáltakNicotiana benthamiana növényeket. Négyféle génkonstrukciót állítottak elő:teljes hosszúságú (full length) CP gén, két csonkított CP gén (az egyikben az N-terminális régióból 46 aminosavat kódoló génszakaszt vágtak ki „NT 138”, míga másikban a konzervatív „core” régióból 86 aminosavat kódoló szakaszttöröltek „CT 258”) és egy antiszensz CP gén konstrukciót. A teljes hosszúságúés az NT 138 transzgéneket hordozó növényekből kimutatták a mRNS-t és afehérjeterméket, míg a CT 258 és az antiszensz konstrukciókkal transzformáltvonalakból csak a mRNS-t tudták kimutatni, a fehérjét nem. PAMV-vel történőfertőzést követően a tünetek kialakulásának késését tapasztalták a teljeshosszúságú, a CT 258 és az antiszensz géneket hordozó vonalakban. Az NT 138konstrukciót hordozó növényekben azonban nem alakult ki ellenállóképesség.A leghatékonyabbnak az antiszensz géneket hordozó konstrukciók bizonyultak.A csonkított „core” régió felhasználásával kimutatták, hogy CP általirezisztencia kiváltásához nemcsak a teljes hosszúságú géneket hordozókonstrukciókat lehet felhasználni.
Chia és mtsai (1992) cymbidium mozaik vírus (cymbidium mosaicpotexvirus –CyMV) CP génjével transzformáltak dohánynövényeket, melyekrezisztensek voltak a későbbi CyMV fertőzésekkel szemben.
Hoekema és mtsai (1988) PVX CP génnel transzformáltakburgonyanövényeket, és 1 µg/ml PVX koncentrációval történő fertőzés esetén

116
a tünetkifejlődés késését és vírusakkumuláció-csökkenést tapasztaltak atranszgénikus vonalakban.
Jongedijk és mtsai (1992) Bintje és Escort burgonyafajtákattranszformáltak szintén PVX köpenyfehérje génnel. A szántóföldi vizsgálatoksorán pozitív korrelációt tapasztaltak az akkumulált CP szint és a rezisztenciaszintje között, azonban ez a korreláció a közepes szinten expresszálóknál nemvolt egyértelműen kimutatható.
PVX-szel szembeni ellenállóságot váltottak ki van der Wilk és mtsai(1991) dohánynövényekben, míg Lawson és mtsai (1990) Russet Burbankburgonyában. Ebben az esetben a transzgénikus növény fogékony volt PVY-nalszemben, azonban egy olyan konstrukció használata által, mely nemcsak aPVX, hanem a PVY köpenyfehérje génjét is tartalmazta, a növények ellenállóváváltak mindkét vírussal szemben. Fehér és mtsai (1992) a PVX 'G'izolátumának CP génjét használták fel burgonya transzformációkhoz. Desiréeés Gracia gumókorongokat transzformáltak Agrobacterium segítségével. A CPgén expressziós szintje alacsony, de kimutatható volt (Northern, Westernanalízissel). Az inokulált levelekben a kontrollokhoz képest a vírus RNScsökkent akkumulációját tapasztalták a transzformánsok nagy részében, sőt 18nappal a fertőzés után egy vonalban 4-10-szeres csökkenést észleltek.
Potyvirus nemzetség
(lásd a dolgozatban, 25. old)
Tenuivirus nemzetség
Rizsprotoplasztok elektroporációs transzformációjához Hayakawa és mtsai(1992) a rizs csíkosság vírus (rice stripe tenuivirus –RSV) köpenyfehérje génjéthasználták fel, és ennek következtében RSV fertőzéssel szemben ellenállónövényeket kaptak. Minden egyes kalluszvonalból számos növénytregeneráltak. A transzgénikus növények magas szinten (0.5 %-ban)expresszálták a CP-t az összes oldható fehérjéhez viszonyítva. Arezisztenciavizsgálatokat az utódnemzedékeken végezték. A fertőzéstsarkantyús kabócákkal (Delphacidae család) végezték. Nyolc héttel a fertőzéstkövetően a transzgénikus növényekben nem volt kimutatható a vírus általkódolt protein, ami arra utalt, hogy a védekezőmechanizmus a vírusreplikációelőtt lépett fel. A nem-transzgénikusok 80 %-a, valamint a CP-t nem termelőtranszgénikusok azonban 15 nappal a fertőzés után megbetegedtek. Ez a kísérletbizonyította, hogy a köpenyfehérje által indukált rezisztencia jelensége

117
kiterjeszthető a gabonafélékre is és olyan vírusok esetében is létrejöhet,amelyek csak rovarvektorokkal terjednek.
Tobamovirus nemzetség
Paradicsom transzformációhoz használták fel Sanders és mtsai (1992)valamint Nelson és mtsai (1988) a TMV 'U1' törzs illetve a paradicsom mozaikvírus (tomato mosaic tobamovirus –ToMV) CP génjét. A TMV CP-veltranszgénikus növények rezisztensek voltak az 'U1' és egy súlyosabb károkatokozó, un. 'PV 230'-as törzzsel szemben, viszont ToMV fertőzés eseténfogékonyként viselkedtek. A ToMV köpenyfehérjét hordozó vonalak védettekvoltak ToMV fertőzéssel szemben. A CP-t alacsonyabb szinten expresszálóvonalak erősebb rezisztenciával rendelkeztek, mint a magasabb szintenexpresszálók. A TMV CP és a ToMV CP-k közötti homológia eléri a 88 %-ot,ez azonban a korábban tapasztaltaktól függetlenül, mégsem eredményezettellenállóságot a TMV CP - ToMV fertőzéssel szemben.
Clark és mtsai (1990) dohánynövényeket transzformáltak TMV CP-ttartalmazó konstrukciókkal. Egy esetben promóterként a 35 S promóterthasználták, más konstrukciókban pedig az rbcS promótert, mely elsődlegesen amezofillumsejtekben működik. A vizsgált transzgénikus növényvonalak eltérőrezisztenciával rendelkeztek, melyet a felhasznált promóter kimutathatóanbefolyásolt. A 35S esetében az ellenállóképesség a növény egészében erősebbvolt, kivéve a mezofillumsejteket illetve a protoplasztokat, ahol a két különbözőpromóter használata esetén nem találtak különbséget a rezisztencia szintjeközött.
Reimann-Philip és Beachy (1993) kimutatták, hogy a TMV köpenyfehérjegénnel transzformált dohánynövényekben a rezisztencia kialakításáhozszükséges a CP jelenléte. Tisztított vírus RNS-sel történő fertőzésnél azellenállóképesség erőteljesen lecsökkent, de teljesen nem tűnt el. Aköpenyfehérjét felhalmozó vonalak nem fertőződtek meg, míg azok, amelyeknem halmozták fel a CP-t alacsonyabb víruskoncentrációnál is fertőződtek. Aköpenyfehérje termelődése nemcsak TMV-vel szemben eredményezettrezisztenciát, hanem más, a tobamovírus nemzetségbe tartozó rokon törzsekkelszemben is, ez viszont függött a CP-k aminosavhomológiájától. Feltételezték,hogy a köpenyfehérje valószínűleg az RNS 5' végére kötődve gátolja ariboszómákhoz kötődését. A CP által kiváltott rezisztencia TMV esetében eltéra klasszikus keresztvédettség jellemzőitől, mivel abban az esetben közvetlenkapcsolata van a fertőző vírusnak a gyengített vírus köpenyfehérjéjével, illetveRNS-ével vagy valamely növényi védekezési reakcióval, míg CPMR esetében a

118
dohány és a TMV kapcsolatban nem találtak speciális vagy általános növényiválaszreakciót.
Clark és mtsai (1995) is azt tapasztalták, hogy transzgénikusdohánynövények, melyek a TMV 'U1' törzsének köpenyfehérje génjétexpresszálták rezisztensek voltak több tobamovírussal szemben is. A fertőzővírus felületi fehérjéi szerepének vizsgálatára mutáns TMV 'U1' konstrukcióthoztak létre, mely a bengáliai kender vagy Crotalaria mozaik vírus (sunn hempmosaic tobamovirus –SHMV) amino- vagy karboxiterminális régiójáttartalmazta. A mutáns vírus nem törte át a TMV által előidézett rezisztenciát,viszont az a kiméra konstrukció, melynek köpenyfehérje génje teljes mértékbenaz SHMV-től eredt, az SHMV-hez hasonló tüneteket idézett elő. Ez azeredmény az ellenállóság kialakításának szempontjából a transzgéntermék és afertőző vírus aminosavszekvenciái közötti interakcióra utal, melybenvalószínűleg nem a köpenyfehérje felületén található aminosavak játszanak főszerepet.
Tobravirus nemzetség
Van Dun és mtsai (1987), illetve van Dun és Bol (1988) dohány rattle vírus(tobacco rattle virus –TRV) 'TCM' törzsének köpenyfehérje génjét használtákfel Samsun dohánynövények transzformációjához. A cDNS a CP-n kívültartalmazott szekvenciákat az 5' nemkódoló és a 3' nemkódoló régiókból is,több, mint 400 nukleotidot. Ezt a 35 S promóter és a NOS terminátor közéépítették be. A „CP+” vonalakban 0.05 %-ban mutatták ki a köpenyfehérjét azösszfehérjéhez viszonyítva. Ezekben a vonalakban enyhe nekrózist figyeltekmeg 1 µg/ml TRV 'TCM' fertőzést követően, viszont nem tudták kimutatni aTRV RNS-t. Ezzel szemben a „CP-” növényekben nekrotikus foltok jelentekmeg az inokulált leveleken és súlyos nekrózis a szisztemikusan fertőződöttszáron és leveleken. A „CP+” vonalakat további vizsgálatokba bevonvafertőzték TRV 'PLB' törzzsel valamint borsó korai barnulás vírussal (pea earlybrowning tobravirus –PEBV). A TRV 'TCM' és 'PLB' törzseinekköpenyfehérjéi közötti aminosavszekvencia-homológia megközelítőleg 39 % ésmindkét CP képes a másik törzs RNS-ének becsomagolására. A PEBV izolátumés a 'TCM' törzs köpenyfehérjéi közötti homológia ennél alacsonyabb szintű,ezért meglepő, hogy míg a „CP+” növények TRV 'PLB' fertőzéssel szembenérzékenyek voltak, addig PEBV-vel szemben rezisztensnek bizonyultak.Mindebből arra következtettek, hogy valószínűleg nem az RNS:CP interakció aköpenyfehérje által kiváltott rezisztencia fő hatásmechanizmusa, hiszen a kétTRV képes a másik RNS-t becsomagolni, viszont védelmet nem nyújtottakegymással szemben.

119
Angenent és mtsai (1990) pedig TRV 'PLB' izolátum CP génjéveltranszformáltak dohánynövényeket, melyek TRV 'PLB' fertőzéssel szembenrezisztensek voltak, TRV 'TCM' fertőzés esetén azonban fogékonyak.
Tombusvirus nemzetség
Dohánynövények transzformációjára használták fel a cymbídiumgyűrűsfoltosság vírus (cymbidium ringspot tombusvirus –CymRSV) CP génjétRubino és mtsai (1993). A N. benthamiana növények csupán 0.05 µg/mlCymRSV inokulumkoncentráció esetén voltak védettek. A rezisztencia szintjeaz alacsonytól a közepesig terjedt. Magasabb, azaz 0.5 vagy 5 µg/mlvíruskoncentráció esetén csupán néhány napos tünetkifejlődés-késésttapasztaltak. RNS-sel történő fertőzést követően pedig a növények fogékonyakvoltak.
Tospovirus nemzetség
De Haan és mtsai (1992) valamint MacKenzie és Ellis (1992) a paradicsomfoltoshervadás vírus (tomato spotted wilt tospovirus –TSWV) N génjéthasználták fel dohánynövények transzformációjára. A vírus negatív szálú RNS-ttartalmaz, melynek N génje képezi a nukleokapszidot. TSWV-vel történőfertőzést követően magas szintű rezisztenciát figyeltek meg tripsz átvitel eseténis, amely azonban más tospovírusokkal szemben nem működött, holott akár 80%-os homológia is előfordul az egyes vírusok között. 23 vonalból 4-bentranszlációdefektív transzgén volt, de ezekben szintén nagyfokú rezisztenciáttapasztaltak. Megfigyelték, hogy a rezisztencia a nem-transzlálódott N gén RNSakkumulációjának függvényében változott illetve függött a transzgén és afertőző vírusgén közötti homológia mértékétől.
Goldbach és de Haan (1993) szintén TSWV N génnel transzformáltaknövényeket, melyek tripsz általi fertőzéssel szemben is rezisztensek voltak. Atranszlációdefektív transzgéneket hordozó vonalak is ellenállónak bizonyultak,mely eredmények az előzőekkel egybevágóan a rezisztencia RNS-függőségétbizonyították.
Pang és mtsai (1993) TSWV 'BL' szensz és antiszensz nemtranszlálódó Ngénekkel transzformáltak dohánynövényeket, melyek rezisztensek voltakhomológ és közeli rokon izolátumokkal szemben, azonban távolabbi rokontospovírusok esetében nem. Az így létrejövő RNS-függő rezisztencia azoknál avonalaknál volt a leghatékonyabb, melyek alacsony szinten expresszálták azRNS-t. Protoplasztkísérletekből levont következtetések alapján feltételezték,hogy az ellenállóság a replikáció gátlásából ered. Ellentétben a szensz RNS

120
általi rezisztenciával, az antiszensz RNS-ek esetében a védelmi reakció függöttaz inokulumkoncentrációtól és a növény korától. Korábbi tapasztalatokhozhasonlóan a homológ és a közeli rokon vírusokkal szembeni rezisztenciaesetében alacsony szintű volt az N gén expressziója és nem volt kimutatható afehérjetermék.
Vaira és mtsai (1995) TSWV egyik olasz izolátumának N génjéveltranszformáltak N. benthamiana növényeket. A transzgénikus vonalakattesztelték TSWV izolátumokkal és 3 másik tospovírussal szemben:földimogyoró rügynekrózis vírus (groundnut bud necrosis tospovirus –GBNV),földimogyoró gyűrűsfoltosság vírus (groundnut ringspot tospovirus –GRSV),nebántsvirág nekrotikusfoltosság vírus (impatiens necrotic spot tospovirus –INSV). Tizenhárom vizsgált vonalból kettő bizonyult rezisztensnek a többiTSWV izolátummal szemben, azonban ezekben a növényekben a transzgénexpresszió alacsonyabb volt, mint bármelyik fogékony vonalban, és az Nprotein hiányzott vagy nem volt detektálható. Más tospovírusokkal történőfertőzésekkel szemben viszont fogékonyak voltak, egyedül GRSV esetébentapasztaltak enyhébb szimptómákat. Azokban a vonalakban, melyek nagyobbmennyiségű proteint termeltek, 2-3 hetes késéssel fejlődtek ki a fertőzésre utalótünetek legalább egy TSWV izolátummal szemben. Szintén késést vagygyengülést észleltek INSV és/vagy GRSV által okozott tünetekben.Megfigyelték tehát, hogy a magas N gén expresszió esetenként visszatarthatja aszimptómák kifejlődését TSWV és INSV-vel történő fertőzésnél, azonban adetektálható expresszió hiánya TSWV-specifikus rezisztenciát eredményez.Pang és mtsai (1993) kimutatták, hogy a TSWV rezisztencia alacsony szintenexpresszálóknál RNS-függő, míg magas szinten expresszálóknál TSWV-vel ésINSV-vel szemben protein-függő. Gielen és mtsai (1991) azonban nem találtakösszefüggést a rezisztencia szintje és a fehérjetermék mennyisége között.Feltételezték, hogy a védettség RNS-függő. GRSV esetében magas szintenexpresszálóknál tünetkifejlődés -késést tapasztaltak, míg a korábbi kutatásieredmények nem találtak összefüggést az expressziós szint és a rezisztenciaközött ennél a vírusnál. GBNV-vel szemben minden vonal fogékonynakbizonyult, ami valójában nem meglepő, mivel genetikailag távolabbhelyezkedik el a TSWV-től.
Prins és mtsai (1995) transzgénikus dohánynövényeket állítottak elő olyangénkonstrukcióval, mely 3 tospovírus köpenyfehérje génjeit tartalmazta 35 Spromóter és NOS terminációs szignálok közé építve. A TSWV, a GRSV és aparadicsom klorotikus foltosság vírus (tomato chlorotic spot tospovirus –TCSV) köpenyfehérje géneket hordozó transzgénikus vonalak magas fokúrezisztenciával rendelkeztek mindhárom vírussal szemben, ezáltal lehetőségetadva széles spektrumú rezisztencia kialakítására.

121
Burgyán és mtsai (1998) TSWV 'Hm' izolátumának nukleoprotein/N génjétépítették be Agrobacterium segítségével dohánynövényekbe. 82 függetlentranszfománs vonalat kaptak, melyek közül 8 bizonyult vírusellenállónak. Ezeka vonalak az ismételt felülfertőzéssel szemben is ellenállóak voltak. A tesztelésTSWV 'Hm' izolátummal fertőzött dohánynövények szövetnedvével,mechanikai úton történt. 14 nappal ezt követően a növények fele atranszgénmentes kontrollokhoz hasonló tüneteket mutatott (erős szisztemikusmozaik, levéldeformáció, nekrotikus lokális léziók). Majd ezek a növényeklassú ütemben el is pusztultak. 13 tünetmentes növényt újra inokuláltak,amelyekből 5 vonal esetében két héttel később még nem, de egy hónapelteltével már lokális léziókat figyeltek meg, majd ezek egyre súlyosabbtünetekké fejlődtek. Így ezek vírusellenállósága részleges volt, míg atünetmentes 8 növényen két hónap elteltével sem alakultak ki a tünetek. Avírust is csupán a tünetes növényekből tudták kimutatni.

122
2. MellékletKöpenyfehérje gének által kiváltott rezisztencia vírusnemzetségenként
Vírus-
nemzetség
beépített
CP
transzformált
növényrezisztencia referencia
AMVdohány,
paradicsomAMV (+)
Tumer és mtsai, 1987,
1991
AMV-
YSMVdohány
AMV-YSMV (+),
TSV (-)van Dun és mtsai, 1987
AMV-425 dohány, lucernaAMV-425 (+),
AMV-McKinney (+)
Loesch-Fries és mtsai,
1987
AMV lucerna AMV (+) Halk és mtsai, 1989
AMV dohányPVX (+), CMV (+-),
TMV(-)
Anderson és mtsai,
1989
AMV dohány, lucerna AMV (+) Hill és mtsai, 1991
Alfamo-virus
AMVdohány
(Xanthi)AMV (+)
Yushibow és mtsai,
1995
PVS-An N. debneyii PVS-ME (+)MacKenzie és
Tremaine, 1990
Carla-virus
PVS-An
burgonya
(Russet
Burbank)
PVS-An (+),
PVM (+-)
MacKenzie és mtsai,
1991
CMV-C dohány CMV-C (+) Cuozzo és mtsai, 1988
CMV-Cdohány
(Xanthi)
CMV-C (+), CMV-
Chi(+),
CMV-WL(+-)
Quemada és mtsai,
1991
CMV-WL dohány
CMV-WL (+), CMV-C
(+),
CMV-Chi (+)
Namba és mtsai, 1991
CMV tök CMV (+)Gonsalves és mtsai,
1992
CMV-O dohányCMV-O (+),
CMV-Y (+)
Nakajima és mtsai,
1993
Cucumo-
virus
CMV-Y dohány CMV-Y (+) Okuno és mtsai, 1993
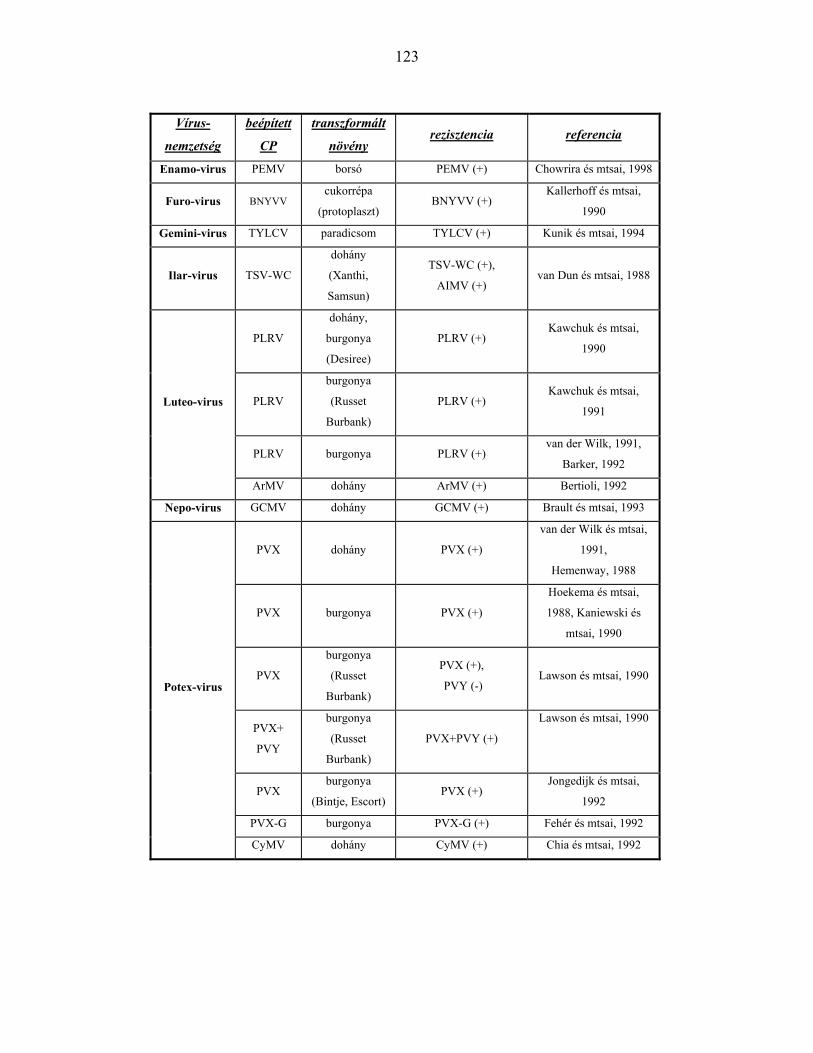
123
Vírus-
nemzetség
beépített
CP
transzformált
növényrezisztencia referencia
Enamo-virus PEMV borsó PEMV (+) Chowrira és mtsai, 1998
Furo-virus BNYVVcukorrépa
(protoplaszt)BNYVV (+)
Kallerhoff és mtsai,
1990
Gemini-virus TYLCV paradicsom TYLCV (+) Kunik és mtsai, 1994
Ilar-virus TSV-WC
dohány
(Xanthi,
Samsun)
TSV-WC (+),
AIMV (+)van Dun és mtsai, 1988
PLRV
dohány,
burgonya
(Desiree)
PLRV (+)Kawchuk és mtsai,
1990
PLRV
burgonya
(Russet
Burbank)
PLRV (+)Kawchuk és mtsai,
1991
PLRV burgonya PLRV (+)van der Wilk, 1991,
Barker, 1992
Luteo-virus
ArMV dohány ArMV (+) Bertioli, 1992
Nepo-virus GCMV dohány GCMV (+) Brault és mtsai, 1993
PVX dohány PVX (+)
van der Wilk és mtsai,
1991,
Hemenway, 1988
PVX burgonya PVX (+)
Hoekema és mtsai,
1988, Kaniewski és
mtsai, 1990
PVX
burgonya
(Russet
Burbank)
PVX (+),
PVY (-)Lawson és mtsai, 1990
PVX+
PVY
burgonya
(Russet
Burbank)
PVX+PVY (+)
Lawson és mtsai, 1990
PVXburgonya
(Bintje, Escort)PVX (+)
Jongedijk és mtsai,
1992
PVX-G burgonya PVX-G (+) Fehér és mtsai, 1992
Potex-virus
CyMV dohány CyMV (+) Chia és mtsai, 1992
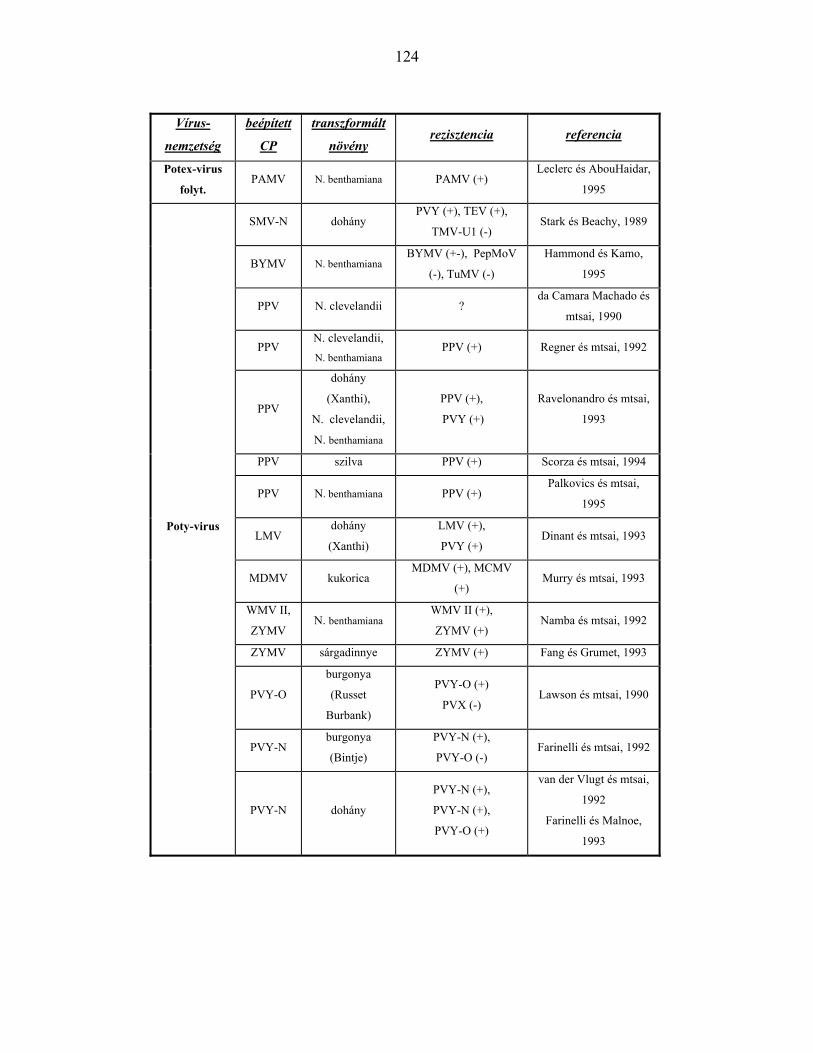
124
Vírus-
nemzetség
beépített
CP
transzformált
növényrezisztencia referencia
Potex-virus
folyt.PAMV N. benthamiana PAMV (+)
Leclerc és AbouHaidar,
1995
SMV-N dohányPVY (+), TEV (+),
TMV-U1 (-)Stark és Beachy, 1989
BYMV N. benthamianaBYMV (+-), PepMoV
(-), TuMV (-)
Hammond és Kamo,
1995
PPV N. clevelandii ?da Camara Machado és
mtsai, 1990
PPVN. clevelandii,N. benthamiana
PPV (+) Regner és mtsai, 1992
PPV
dohány
(Xanthi),
N. clevelandii,
N. benthamiana
PPV (+),
PVY (+)
Ravelonandro és mtsai,
1993
PPV szilva PPV (+) Scorza és mtsai, 1994
PPV N. benthamiana PPV (+)Palkovics és mtsai,
1995
LMVdohány
(Xanthi)
LMV (+),
PVY (+)Dinant és mtsai, 1993
MDMV kukoricaMDMV (+), MCMV
(+)Murry és mtsai, 1993
WMV II,
ZYMVN. benthamiana
WMV II (+),
ZYMV (+)Namba és mtsai, 1992
ZYMV sárgadinnye ZYMV (+) Fang és Grumet, 1993
PVY-O
burgonya
(Russet
Burbank)
PVY-O (+)
PVX (-)Lawson és mtsai, 1990
PVY-Nburgonya
(Bintje)
PVY-N (+),
PVY-O (-)Farinelli és mtsai, 1992
Poty-virus
PVY-N dohány
PVY-N (+),
PVY-N (+),
PVY-O (+)
van der Vlugt és mtsai,
1992
Farinelli és Malnoe,
1993

125
Vírus-
nemzetség
beépített
CP
transzformált
növényrezisztencia referencia
PVY dohány PVY-NTN (+) Kollár és mtsai, 1993
PVY burgonya PVY (+)
Smith és mtsai, 1995,
Stark és Beachy, 1989,
Kaniewski és mtsai,
1990
TVMV dohányTVMV (+),
TEV (+)Murphy és mtsai, 1990
PRSV dohány, papaya
PVY, PepMoV, TEV
(+),
CMV-C(-)
Ling és mtsai, 1991
PRV papaya PRV (+) Fitch és mtsai, 1992
PStV N. benthamianaPStV (+),
TEV (-)
Cassidy és Nelson,
1995
TEV dohány TEV (+-)Lindbo és Dougherty,
1992b
Poty-virus
folyt.
TEVdohány
(Burley 49)TEV (+)
Dougherty és mtsai,
1994
Tenui-virus RSV rizs RSV (+)Hayakawa és mtsai,
1992
TMV-U1 dohány TMV-U1 (+) Powell és mtsai, 1986
TMV-U1 paradicsom
TMV-U1, TMV-PV230
(+),
ToMV-L (+), ToMV-2
(+-)
Nelson és mtsai, 1988
TMV dohány
TMV, ToMV,
TMGMV, ORSV,
PMMV (+),
RMV, SHMV (-)
Nejidat és mtsai, 1990
Tobamo-
virus
TMV dohány TMV (+)
Anderson és mtsai,
1989, Clark és mtsai,
1990, Reimann-Philip
és Beachy, 1993

126
Vírus-
nemzetség
beépített
CP
transzformált
növényrezisztencia referencia
TMV-U1 dohány mut. TMV (+),
TMV-U1+SHMV CP
(-)
Clark és mtsai, 1995
Tobamo-
virus folyt.ToMV-C paradicsom TMV-U1 (+),
ToMV-C (+)
Sanders és mtsai, 1992
TRV-
TCM
dohány TRV-TCM (+) van Dun és mtsai, 1987
TRV-
TCM
dohány
(Samsun)
TRV-TCM (+), PEBV
(+),
TRV-PLB(-)
van Dun és Bol, 1988
Tobra-virus
TRV-PLB dohány TRV-PLB (+),
TRV-TCM (-)
Angenent és mtsai,
1990
Tombus-
virus
CyRSV dohány CyRSV (+) Rubino és mtsai, 1993
TSWV dohány TSWV (+) MacKenzie és Ellis,
1992, de Haan és mtsai,
1992, Goldbach és de
Haan, 1993
TSWV-
BL
dohány TSWV-BL (+) Pang és mtsai, 1993
TSWV+T
CSV+GR
SV
dohány TSWV (+),
TCSV (+),
GRSV (+)
Prins és mtsai, 1995
TSWV
olasz iz.
N. benthamiana TSWV, INSV,
GRSV(+-),
GBNV(-)
Gielen és mtsai, 1991
Vaira és mtsai, 1995
Tospo-virus
TSWV-
Hm
dohány TSWV-Hm (+) Burgyán és mtsai, 1998


















