
99 - DTIC · measurement of PLA.1 induced rat red blood cell ghost IRBC-G) hemolysis. PlA.', was...
15
bnflaniunaion. Vol. 13, No. 1, 90 M R3 1980 ,. MODULATION OF PHOSPH-OLIPASE A, LYTIC 1.O ACTIVITY BY ACTIN AND MYOSIN' D. A. DUBOSE, 2 D. SHEPRO.' and H. B. HECHTMAN 4 (D 2 ~US Arjiv Rc'scarc/i Initw ilifEnc]1vironnienccl Medicine 0 Natick. Massachusc'irs 01 760 'Department of Biologv/.Surgerv, Boston Universiity 'H-arvard Medical School. Boston, Mu.scc/us'as 02215 0Ahstrac 7 ostacsclin IPGI> ) production is closely coupled with endothelial cell shape and F-actin distribution in vitro. These findings may implicate c\toskeletal constituents in a mechanism regulating eicosanoid metabolism. To determine the potential for such a regulatory mechanism. cytoskeletal protein effects on the rate- limiting cicosanoid cascade cnzinme (phospholipase A 2 PLA') were studied. Memi- brane phospholipid dJgradation was indirectly determined by spectrophotomectric measurement of PLA.1 induced rat red blood cell ghost IRBC-G) hemolysis. PlA.', was incubated 'A.ith- j in (skeletal, smooth, or nonmusele cell) at a nonmruscle cell concentrat ion 000q0 .l and then exposed to the RBC-G. Comparisons in the pres- encc5 r absence of actin revealed that F-actin stimulated whereas 0-actin suppressed PLA 2 lytic behavior significantly (P < 0.1)5). When a 10: or 100i I F-aein to myosin ratio %vas used. the F-actin stimulator-s effect was significantly (P < (0.0)5) reduiccd. These findings suggest that the in vitro correlation between PC;I~ product ion and endothelial cell shape ma, ,be the result of PLA7'regulation bN cvtoskectal elements, that impart cellular form. >Thr. '. ( INTRODUCTION Prostacyclin (PGI,) affects the cardiovascular system in numerous ways (1). one of' which is augmentation of actin stress fiber formation (2). These are cytoskeletal elements that in vitro appear to promnote endothelial cel Junctional integrity (3). Thus, this eicosanoid may influence permeability, and knowledge The investigators adhered to the "Guide for Laboratory Animal Facilities and Care" as promul- gatedf by the commiiiittee on the Gutide for Labi rators Animal Faciities andi Care of' the Institute oft Laborators Amnal Resources. National Academyv oft Sciences. National Research Council. The opinons or assertions contained herein are the private views of'the authors and are not to be construed as oftficial or as reflecting the % e%% s of the Decpartmoent tif the Arn\ ior the D)epartment ot Defense. .1; I 1u i 1111 l 4- uhD.nA 99 ~ ~ w%
Transcript of 99 - DTIC · measurement of PLA.1 induced rat red blood cell ghost IRBC-G) hemolysis. PlA.', was...

bnflaniunaion. Vol. 13, No. 1, 90 M R3 1980 ,.
MODULATION OF PHOSPH-OLIPASE A, LYTIC1.O ACTIVITY BY ACTIN AND MYOSIN'
D. A. DUBOSE,2 D. SHEPRO.' and H. B. HECHTMAN4
(D 2 ~US Arjiv Rc'scarc/i Initw ilifEnc]1vironnienccl Medicine
0 Natick. Massachusc'irs 01 760'Department of Biologv/.Surgerv, Boston Universiity
'H-arvard Medical School. Boston, Mu.scc/us'as 02215
0Ahstrac7 ostacsclin IPGI> ) production is closely coupled with endothelial cellshape and F-actin distribution in vitro. These findings may implicate c\toskeletalconstituents in a mechanism regulating eicosanoid metabolism. To determine thepotential for such a regulatory mechanism. cytoskeletal protein effects on the rate-limiting cicosanoid cascade cnzinme (phospholipase A2 PLA') were studied. Memi-brane phospholipid dJgradation was indirectly determined by spectrophotomectricmeasurement of PLA.1 induced rat red blood cell ghost IRBC-G) hemolysis. PlA.',was incubated 'A.ith- j in (skeletal, smooth, or nonmusele cell) at a nonmruscle cellconcentrat ion 000q0 .l and then exposed to the RBC-G. Comparisons in the pres-encc5 r absence of actin revealed that F-actin stimulated whereas 0-actin suppressedPLA2 lytic behavior significantly (P < 0.1)5). When a 10: or 100i I F-aein to myosinratio %vas used. the F-actin stimulator-s effect was significantly (P < (0.0)5) reduiccd.These findings suggest that the in vitro correlation between PC;I~ product ion andendothelial cell shape ma, ,be the result of PLA7'regulation bN cvtoskectal elements,that impart cellular form. >Thr. '. (
INTRODUCTION
Prostacyclin (PGI,) affects the cardiovascular system in numerous ways (1).one of' which is augmentation of actin stress fiber formation (2). These arecytoskeletal elements that in vitro appear to promnote endothelial cel Junctionalintegrity (3). Thus, this eicosanoid may influence permeability, and knowledge
The investigators adhered to the "Guide for Laboratory Animal Facilities and Care" as promul-gatedf by the commiiiittee on the Gutide for Labi rators Animal Faciities andi Care of' the Instituteoft Laborators Amnal Resources. National Academyv oft Sciences. National Research Council.The opinons or assertions contained herein are the private views of'the authors and are not to beconstrued as oftficial or as reflecting the % e%% s of the Decpartmoent tif the Arn\ ior the D)epartmentot Defense.
.1; I 1u i 1111 l 4- uhD.nA
99 ~ ~ w%

16 i)uBose et al.
of the mechanisms that control its production might provide insight to altera-tions in endothelial cell barrier function.
The rate-limiting enzyme of the PGI2 cascade is phospholipase A, (PLA,)(4). This enzyme is soluble or membrane associated (5) and may be influencedby the modulatory effects of calcium, cAMP, hormones, and membrane struc-ture (6). Cell shape is perhaps another regulatory factor. For example, brady-kinin stimulates PG12 production by substrate-attached endothelial cells (7). Incontrast. endothelial cells (8) or transformed fibroblasts (7) in suspension arenot stimulated by bradykinin. The cell shape differences of attached and sus-pended cells perhaps explain this phenomenon. Moreover, PGI2 synthesis bythe small endothelia! cells in cotact-inhibiled cultures might exceed that oflarge cells in nonconfluent cultures because of cell shape differences (9). Cellshape is of greater significance than cell-to-cell contact, since in the absence ofcontact inhibition, small endothelial cells still generate more PGI, than largecells (10). Small endothelial cells in culture have a diffuse F-actin distributionand lack the F-actin stress fibers seen in large cells (Figure 1). Enhanced PGI,synthesis occurs with a diffuse F-actin distribution (Figure IA) and productionis reduced when complexes of F-actin, myosin, and other cytoskeletal proteins
(stress fibers. Fig'ire IB) are present (10).Changes in the distribution and polymerized state of actin alter cell shape.
Moreover. actin binds many proteins (11) and regulates glvcolysis by allostericmodulation of glycolytic enzyme catalytic site (12-15). Thus. cytoskeletal fac-tors that influence cell shape are also associated with metabolic control mech-anisms. Because cell shape (7-10) and F-actin distribution (10) correlate withPGI production, PLA, regulation by a cytoskeletal-mediated mechanism maybe operational in eicosanoid metabolism. To determine the potential for such a
mechanism, the effect of actin and actin-myosin complexes on extracellularPLA, lytic behavior was determined. These cytoskeletal elements were asso-ciated with an alteration in PLA, lytic activity, which further suggests the con-trol of intracellular PLA, by the constituents that impart cellular form.
MATERIALS AND METHODS
Calcium chloride, ATP. potassium chloride. guanidine hydrochloride. rabbit muscle atin.chicken muscle actin. chicken gizzard actin. bovine muscle myosin. chicken git,ard mi~osin. bo incalbumin, phalloidin. and PLA. derived f'ronm bee venom (PLA,-B )or porcine pancreas (P.A.-Piwere obtained from Sigma Chemical Co.. St. Louis, Missouri. Human platelet actin i.as procured1nm Calbiochem Biochemicals. San Diego, California. Sodium chloride and sodium acetate w.ere
purchased front Mallinckrodt, Paris. Kentucky. Fisher Scientific iFairlawn. Ncvs Jersey) A~as thesource for Tris and EDTA.
The compositions of bu tiers and solutions %%ere as lblos s: isotonic saline, I. 144 M sodiuii

Actin and Myosin PLA 2 Regulation 17
Fig. I. The effects of substrate adhesive capacity on bovine aortic endothelial (BAE) cell shape.F-actin distribution, and actin stress fiber formation. BAE cells were seeded on bacteriologicalplastic or tissue culture plastic. After 24 h. cells were fixed (3.7% foninalin, 30 nini. treated A ithacetone (8(0e/, 10 rin), and exposed to rhodamine phalloidin (0. 165 pM, 10 min, ]unction Cits.Oregon). A Zeiss microscope equipped with the appropriate filters for the excitation o" rhodaninewas used to observe the cell fluorescence. Photographs of the fluorescence were taken using I ri-XPan film (Kodak, 4(X) ASA). BAE cells on a substrate of reduced adhesi.e capacit% (bacteriologicalplastic. A) were small with a difff's'e F-actin distribtion :rd appeared to lack stress fibers. Suchcells on an adhesive substrate (tissue culture plastic. B) were large \4 ith man% discrete actin stressfibers. Bar equals 10 lpt.
(,!...%mmm im iimmmmmmv mam m

18 I)uBose et al.
chloride. 2.0 mM calcium chloride. pH 7.5, hemolysis buffer, 0.05 M sodium chloride. 1.0 mMEDTA: resealing solution. 1.5 M sodium chloride, 1.0 mM EDTA; actin polymerizing solution.50.0 mM potassium chloride, 2.0 mM calcium chloride, 1.0 mM ATP; actin depolymerizing solu-tion. 1.5 M guanidine hydrochloride. 1.0 M sodium acetate, 1.0 mM ATP, 20.0 mM Tris. 6.0mM calcium chloride, pH 7.5
Red blood cell ghosts (RBC-G) were prepared as previously described (16. 17). Blood wasobtained from anesthetized (52 mg penobarbital s(diumlkg; intravenous) rats. After centrifugation(2500g) and supematant aspiration. the red blood cells were washed in saline three times. To each5-tnl pellet volume, 50 ml of hemolysis buffer was added and stirred at room temperature tor 30moin. A sufficient volume of rescaling solution was then added to restore isotonicity. This mixturewas stirred slowly for I h. After resealing, ghosts were collected by centrifugation (15,(.OOg). Withgentle resuspension, ghosts were separated from the lysis-resistant red blood cells that comprisedthe dark red button in the pellet center. Ghosts were washed in saline until the absorbance value(418 nm) remained constant. RBC-G were then adjusted to a 20% hematocrit.
Polymerizing solution was employed to generate F-actin (18). Actin (5 mg/ml) was sus-pended in this solution for 2 h at 5°C. Polymerization increased solution viscosity. Actin 15 o3gml) was suspended in depolymerizing solution to obtain G-actin (19). To account for an possibl,:direct polymerizing or depolymerizing buffer effects, these solutions were used as diluents for PLA.incubated in the absence of cytoskeletal protein.
Measurement of PLA. activity via RBC-G lysis followed previously described proccdurcs(16. 17). RBC-G (20% hematocrit) were diluted in isotonic saline ( :5.33) to a 9.0 ml volume(Figure 2). Four 250-al samples were then collected, centrifuged (I rain, 15,OOg), the supcniatalHidiluted (1: 10) in distilled water, and measured in a spectrophotometer at 418 nm to deteminebackground absorbance before PLA, addition. PLA:-B (88 units/mIl) was incubatwd (30 min. 50CIalone or with G- or F-actin ( 100M.M) from bovine or chicken skeletal muscle. Similar PLA.-P 3(X)units/mil) studies employed actin derived from human platelets, chicken skeletal muscle, or chickengizzard. Some PLA_-P studies used incubations with phalloidin (1(X) .M) or bovine albumin (it)
.aM). Other studies involved incubations first with F-actin (chicken skeletal muscle) and then phal-
5 MIN. INTERVALSAMPLES
CENTRIFUGATIONsaline + 2mM CaC, (15.000 xg)
G- OR F- Actin (20% heatoct) 0. su(1001JM) 0.2ml supematant
Myosin (I or 10 pM) 1.00pM Actin +0.01 or 010 pM Myosin 1.8ml HO
PLA2 3.0 u/nol PLA,(300u/mi) Absorbance
418nM
Fig. 2. Phospholipase A. (PLA,) was incubated (3)) in 5'C) in the presence or absence of
cytoskIctal protein prior to exposure to the red blood cell (RB,) ghost bath. Control experimentsconducted in the absence ol V- ot G act/n timed enz mie incubated % ith polneri/ng or depolN,merizing solution, respectively. PL.A2 activity was followed 6s niellu.e ,cit . 'ample sUpcmmtantabsorbance at 418 nm at 5-min inter'als over a 3o-min period.

Actin and Myosin PLA 2 Regulation 19
loidin. PLA.-P was also incubated with F-actin (chicken skeletal muscle or human platelet) andthen myosin (30 min, 5°C. bovine muscle or chicken gizzard. 10 or I pM). After these incubationconditions, an aliquot (80.0 Ml) was added to the RBC-G (8 ml) and gently stirred at 37°C. Over30 min, at 5-mmin intervals, four 250-gl samples were collected and supematant absorbance deter-mined.
Each PLA_ lytic activity study was repeated in triplicate. From the 12 samplings (4 samples.er 5-min time period x 3 replications), a mean absorbance value was calculated and a third-uegree regression generated that described PLA, lytic activity with time. To determine significant(P < 0.05) differences. lytic activity curves were studied by analysis of variance and Tukey com-putation (20).
RESULTS
PLA, action on the RBC-G phospholipid membranes resulted in lysis. Asmeasured by an increased bath solution supernatant absorbance, hemoglobinwas released, which indirectly determined PLA, activity (16, 17). Control stud-ies demonstrated that actin, myosin, or the small volume (80 il) of polymer-izing or depolymerizing solution in the absence of PLA, did not induce ghostlysis (data not shown). Although enzyme activity was not significantly alteredby incubation in polymerizing or depolymerizing solution, activity was signif-icantly enhanced by depolymerizing solution when compared to incubation insaline (Figure 3A). Moreover, differences in the RBC-G from preparation topreparation altered the PLA, kinetics (Figure 3B). To account for these assaycharacteristics, PLA, was incubated with polymerizing or depolymerizing solu-tion in the absence of F- or G-actin, and only activity curves generated fromthe same RBC-G preparation were compared. Under identical experimentalconditions, PLA, lytic activity curves were replicated without significant dif-ferences (Figure 3C). Results were similar using PLA,-B or PLA,-P incubatedin polymerizing or depolymerizing solution.
Significantly, prior incubation of PLA,-B with polymerized chicken or rab-bit muscle actin stimulated, while G-actin reduced, lytic activity when the sameRBC-G preparation was used (Figure 4). Similar actin effects on PLA 2-P activ-ity were noted, even with different RBC-G preparations (Figure 5). For skeletalmuscle (chicken muscle, Figure 5A and B), smooth muscle (chicken gizzard.Figure 5C and D), or nonmuscle cell (human platelet, Figure 5E and F) actin.F-actin significantly stimulated, while G-actin reduced, RBC-G lysis. As illus-trated (Figure 5C and E), prior PLA,-P incubation with F-actin could occa-sionally be associated with an initial suppression in enzyme activity over thefirst 10 min in the RBC-G bath. Incubation with F-actin followed by myosinexposure significantly reduced the stimulation of PLA,-P seen after incubationwith F-actin alone (Figure 6). This was true for both the 100:1 and 10: 1 F-

20 IDuffise et al.
400 - - Polye-- q So'~f-
300
0 5 10 15"
Tine ,,Mmintes
400
PtA * 490-0 P'epo'ot-0n #i
E L PtA . 480-0 P.,P-t-A~ #2
200
10
0
T-me t v4Aes
300
S200-
50-
0 o
0 s
Fig. 3. Effect of different enzyme incubation conditions (saline, polynmerizing, or depols mcrizingsolution in the absence ofcytoskeletal protein) on phospholipase A, (PLA) ltcatt (A). Deni-onstration of the changes in PLA kinetics associated with different ghost preparations (B). Illustra-tion of the close replication of PLA lytic activity curves when the samre enzv,'e source (bee venomnor porcine pancreas). enzyme incubation conditions (polynmerizing or depolymerizing solution inthe absence of cytoskeletal protein), and red blood cell ghost preparation %kere used (C). For A.B. and C. similar results were obtained using PLA derived from bee venom or porcine pancreas.
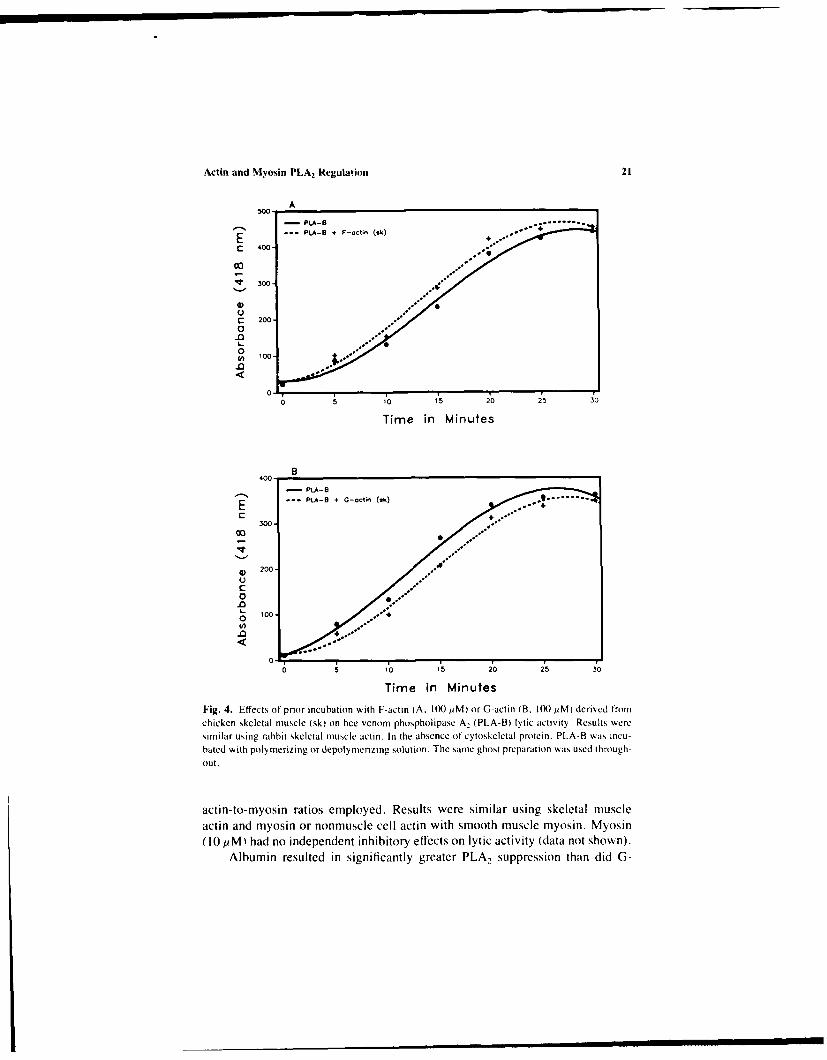
Actin and Myosin PLA2 Regulation 21
A5W0
- Pt.A-8
E ... PLA-8 + F-cctin (ik)
S400-
0
300
.0
Time in Minutes
B400-
.. PLA-B + G-oCtin (2k)
300-
G)200-
000.0
o 5 10 15 20 25 30
rime in MinutesFig. 4. Effects of prior incubation with F-actin (A. 100 1iM) or G-actin (B. 100 pM) derived fromchicken skeletal muscle (sk) on bee venom phospholipase A, (PLA-BI lytic activity. Results weresimilar using rabbit skeletal muscle actin. In the absence tintcytoskeletal protein. PLA-R was incu-hated with poly merizing or depolynIeriZing solution. The samte ghost preparation was used through-out,
actin-to-myosin ratios employed. Results were similar using skeletal muscleactin and myosin or nonmuscle cell actin with smooth muscle myosin. Myosin(10 pM) had no independent inhibitory effects on lytic activity (data not shown).
Albumin resulted in significantly greater PLA, suppression than did G3-

22 DuBose et al.
?-~ 4
I. E E
(wu 9L$,) souoqiosqy (wju Qt,) 03uoqjosq
-4
C --
(wu g9t) a~uoqjoscv (u gs",) aouoqioscv
. a.
-44
cc_

Actin and Myosin PLA2 Regulation 23
350
-PLA-P ...
300- PLA-P +- F-Oct, %
PLA-P + F-octn + Myosln (100 1) %
E - PLA-P + F-octin + Myos. (10) .°
250 ...'
200.
En
0 5 10 15 20 25 30
Time in Minutes
Fig. 6. Effects of prior incubation with F-actin (100 jsM) or F-actin (I00 ,M) plus myosin (10 orI pM) on porcine pancreatic phospholipase A, (PLA-P) lytic activity. Similar results were obtainedusing chicken skeletal muscle actin and such actin (skeletal muscle cell) in combination with bovinemuscle myosin or human platelet actin and such actin (nonmuscle cell) in combination w% ith chickengizzard myosin. In the absence of cytoskeletal protein. PLA-P was incubated with polymerizingsolution. The same ghost preparation was used throughout.
actin (Figure 7A), while phalloidin. another noncytoskeletal protein, had noeffect (Figure 7B). When PLA2-P was first incubated with F-actin (chickenskeletal muscle) and then phalloidin. the stimulatory effect of polymerized actinwas significantly improved (Figure 7C).
DISCUSSION
Studies of endothelial cells in culture demonstrate a significant correlation
among cell shape, F-actin distribution, and PGI, synthesis (7-10). The findingsobtained in this current report support an explanation for this phenomenon basedon the modulation of PLA 2 activity by cytoskeletal constituents that impart cel-
lular form.
RBC-G lysis determined PLA2 activity. !.ysis is associated with phospho-
lipid hydrolysis, since in the absence of hydrolysis erythrocytes are not lysed(16), while in ghosts the major phospholipid classes are degraded (21), and
significant phospholipid breakdown correlates with ghost lysis (22). Therefore,
RBC-G lysis is an indirect measure of phospholipid hydrolysis by PLA.

24 i)uBose et al.
250 AP LA-P
.200 PLA-P . 0-oct~n. 2 . PLA-P + Albo-
I00150
.o 100C0
S50
00
5 10 1,5 20 25 i0
Time in Minutes
600
PLA-P500 * PLA-P + Pholloidn
In 400
300
c
0 200-
0100.
0-
051,0 1,5 20 25 2
Time in Minutes
400 C
PLA-Pe
MLA-P . F-Acta A (n)c 300 wPLAP u-t, (ik) PholordIn
200
0
100
051,0 '5 20 25 30
Time in Minutes
Fig. 7. Effects of prior incubation with albumin ( 100 AM) osr G-actin (chicken smototh ttuscle 1(k)AIM) on porcine pancreatic phospholipase A, (PLA-PJ lytic acti% it), (A). In the absence of1 albuminor G-actin. PLA-P was incubated with depolymerizing solution. The aihumin incubation solutionalso contained depolymerizing solution. Effect ot phalloidin exposure ( tX) pMi on enzyme Iunc-tion (B). Incubations in the presence or absence of phalloidin used polymeriiing solution. Dem-onstration of the enhancement effect by the presence of phalloidin (I(X) PM) on chicken skcletalmuscle F-actin (sk: 100 AM) stimulation of PLA-P lytic activity (C). In the presence or absenceof F-actin or F-actin plus phalloidin, all enzyme incubations were conducted in the presence ofpolymerizing solution. Within each study the same RBC-G preparation was used. but differentpreparations were used between studies.

Actin and Myosin PLA2 Regulation 25
Although an indirect determination may have limitations, it was preferable tothose assays that directly measure hydrolysis of an isolated phospholipid, whichis not part of a membrane composed of the many elements that influenceenzyme-membrane interactions. Because this model accounted for some of theadversities associated with the PLA2 active site gaining access to its substratein a complex membrane, it closely approximated the in vivo conditions underwhich intracellular PLA, must function. The use of extracellular PLA, to drawinference about the nature of intracellular PLA, is an approach used by others(6). Moreover, intracellular PLA, molecular weight is similar to PLA,-P and,like the extracellular form. intracellular PLA, may exist as a zymogen (6).Thus, these forms of PLA, may not be unique. and factors that influence oneform may be relevant to the other.
Under identical conditions (same PLA, source, incubation environment,and RBC-G preparation), the assay stability was demonstrated by activity curvereplication without significant differences (Figure 3C). After actin incubation.PLA, from diverse sources w, , stimulated by F-actin, but inhibited by G-actin.This was true using the same RBC-G preparation throughout (Figure 4) or whendifferent preparations were employed (Figure 5), even though such diflrencescould alter the PLA, kinetics (Figure 3B). Any possible effects of the polymer-izing or depolymerizing solution were eliminated by PLA, incubation with thesolution in the absence of cytoskeletal protein. Finally, the dependence of lyticbehavior on actin polymerization state excluded any participation by contami-nants in the actin. since lysis influenced by contaminants should not changewith actin state. Thus. only the effects of actin could account for these changesin PLA, activity.
When incubated with enzyme, actin (5 mg/ml) exceeded the critical con-centr'tion required for polymerization (11). The RBC-G bath conditions werenot as favorable for F-actin (0.05 mg/nil). However, the F-actin state was main-tained for a sufficient time. because lytic activity was enhanced by F-actin. butnot G-actin (Figures 4 and 5). The further enhancement of lytic activitN byphalloidin (Figure 7C) was additional evidence that the F-actin state was essen-tial to lysis stimulation. Phalloidin binds to F-actin in a I: 1 ratio (23), lowversthe critical concentration for polymerization (24. 25). and stabilizes the F-actinstate by reducing monomer dissociation (24).
Occasionally. F-actin initially suppressed lytic activity (Figure 5C and E).This may be related to an actin fiber length that impeded enzyme-membraneinterations. which was overcome by depolynicrization to a more optimal fibersize. The augmentative effect of phalloidin on stimulated PIA, activity with F-actin (Figure 7C) was evidence for dcpolymcrization in the RBC-G bath in theabsence of phalloidin. Thus, fiber length may affect F-actin stimulation of PLA.Since this suppression was not consistently noted, initial actin fiber length inthe RBC-G bath was perhaps an uncontrolled variable.

26 I)uBoe et al.
Actin and myosin are major stress fiber proteins. Myosin had no indepen-dent inhibitory effects (data not shown), but it reduced the F-actin stimulationof lysis (Figure 6). Tropomyosin and oe-actinin stress fiber proteins (10 AM) didnot alter stimulation (data not shown). Thus, if stress fibers have an impact, itis perhaps due to myosin.
Phalloidin, in the absence of F-actin, had no effect (Figure 7B). However.albumin significantly reduced PLA, lytic behavior (Figure 7A). Albumin's effectis indirect, since it alters the form of the substrate presented to PLA. (26). Thegreater inhibition by albumin relative to G-actin (Figure 7A) suggests that theseproteins may influence enzyme activity by different mechanisms. As demon-strated by the phalloidin effect, not all proteins altered lytic activity. This find-ing (Figure 7B) and others (Figure 3C) illustrated how closely lytic curves werereplicated when activity was not changed. Although the actin-associated mod-ulations in PLA, activity may appear small, because the activity curves withineach condition were so closely replicated, these changes were significant.
Although the mechanism of cytoskeletal protein action on PLA, is notknown, it may be relevant to various cell types. since skeletal, smooth, andnonmuscle cell actin isoforms influenced lytic behavior (Figure 5). The rele-vance to endothelial cells was further enhanced by the use of actin (100 AM)and myosin (I pM) concentrations reported for nonmuscle cells (27). Further-more, because diverse extracellular PLA, sources (PLA2-B and PLA.-P) weresimilarly affected by actin, this may be a common PLA2 attribute.
These findings support a potential for cytoskeletal-mediated regulation ofeicosanoid metabolism. A diffuse F-actin distribution in small, substrate-attached endothelial cell:, (Figure IA) permits greater PGI, production (10).because PLA, may be stimulated by F-actin free of other cytoskeletal proteins(Figures 4A and 5AC,E). Reduced PGI, production by large, substrate-attachedcells (10) (Figure IB) might be the result of enzyme suppression by the actin-myosin complexes of stress fibers (Figure 6). Since reduced cell adhesion andF-actin stress fiber level correlate directly (10, 28) (Figure 1), in the absenceof adhesion. G-actin may dominate to perhaps decrease PLA, activity (Figures4B and 5B.DF) and PGI, synthesis in suspended cells (7, 8). Endothelial cellstress fiber promotion by PGI (2) also supports this mechanism. Factors that
induce an actin-myosin dissociation (stress fiber disruption) might enhancePLA. activity. The resulting PGI, would mediate the return of actin-mvosincomplexe, (stress fibers) to perhaps suppress PLA, and further PGI synthesis.Therefore, PGI, and cytoskeletal proteins may function in a reciprocal signalingloop to modulate cell shape and eicosanoid metabolism (Figure 8). Since thecytoskeleton influences junctional integrity (3), if this mechanism were func-tional in vivo. it might play a role in the alteration of permeability within theendothelium.

Act in and NMyosin PLA2 Regulation 27
PGI,* PGIPGIGI
PGII
'5E
C E P
PLPPA
F-ACTI STRESFIER
Fig 8 Ilusratonofa ptetia rcirocl ignlig loptht prhps ritins s mehaisin he eglaton f icoanod etaolsm nd ~t~keeta srucur. Fctos hatcaue disociaio o F-cln nd ivsi (tres ibr dsrpton)ma sim lat te aw-imtig ez~icotthe icoanod cacad. pospohpae A (P.A,. Ths simuatio wold ndue enancd aachidoicacd elas ad leat posacyli (GI) ~nheis T e inin'o PGI. oispttv
membrane ~~~~~~~~~receptorwolintaetetmainocmlxso'Fatn noiniohrcy-
REFERENCRS
I . \% . , R.199 . Posuc~ch inthecariovaculr sste in ealh ad dseas, K Scr'r
A ig. 8.R0 llstatind oH a poHnial eipra sig\,195naolin loott perha auntionsatio af proans-
catnot se-actin and mvosin stress lir isruptin may. str mla. therat-lmitng2n61n9.o
skeleta prins stes . f'.R Bers) to0 a l thecyoseto and cell shapef I , 987 tress hfiers m iht supess

28 DuBose et al.
olism in endlothelial cells: Evidence for phospholipase A and novel Ca2 -independent phos.-
pholipase C. Biochion, Biophy. Acte 917:296-307.5. BLA suhii t. G.. and R. J. Ft~o~k i.H. 1983. Inhibition of phospholipase. Br. Med. Bull. 39:2601-
264.6- V ss Di s BosciH. K 1980. IntracellIula r jli )spholipase A. Bjccchim. Bcccphvs. Ac 1 604:191I -
246.7. Ho-,(;. S. L.. 1981). Etfect ot bradykinin and thrmbin on prostacyclin synthesis in endlothelial
cells from calf and pig aorta and human umbilical cord vein. Thrombi. Re'k. 18:787-795.8. M.w(Is LRi. D. E.. J. D). PI ARSONs. and J. L. Gokiccc. 1978. Localisation and stimulation oit
prostacyclin production in vascular cells. Vature 271:549-551.9. EiLtoR. A.. 1. Vt.oty\svKY. E. H A. R. At /Ntos.. B. B. Wt KSLER. A. Rs%/. and Z. Ft Ks.
1983. Cultured endothelial cells increase their capacity ito synthesize prostacxclin folio\& ingthe fomination of a contact inhibited cell mnonolayer. J. Ccl Pheniol. 114:179-183.
101. Dt.Bosic. D. A.. D. SHEPRO. and H. B. Hli:ctttMAs. 1987. Correlation amiong endothelial cell.shape. F-aetin arrangement. and prostacyclin synthesis. Lifir V,-; 40:447-453.
11. KORN. E. D. 1978. Biucchcmiistrs of actoniyosin-dependent cell motilitN IA revieA 1. Proc.Nall.ad. Sci. U.S.A. 75:588-599.
12. N4.SrF. C. 1984. interactions between glycolytic cnzvites and components of the c~tomna-trix. J. Cell Biol. 992s-25s.
13. WALSH. T. P.. F. M. Ci..NRKt.. and C. J. NM sits. 1977. Mlodification oft the kinetic param-eters of aldolase on binding to the actin-containing tilaments ofimuscle. Bjoc hen,. J. 165:165-.167.
14. WALSH, T. P.. C. J. MIs~t.RS., D. J. MopION. and F. M. Ci..SRKI . 198 1, The reversible bind-
ing of glycolytic enzymes in bovine skeletal miuscle in response to tetanic stimulation. BiccchiptiBiopiivs. Acia 675:29-39.
I5. WAScH. T. P.. D. 3. WINIOR. F. NI. CI...RKI . C. J1. MAIF.stRS. and D. I. MoRbos. 1980.Binding of aldolase to actin containing filaments. Biocwpm. J. 186:89-94.
16. RoFt.oistN. B.. R. F. A. ZAAAt . P. COIt Rtt S. C. B. Wooostcii and L. L. MI. V.\\DEE+NFN. 197 1. Action of pure phospholipase A. and phospholipase C (in human erN ibroc% teN
and ghosts. Biochim,. Bioplivs. Acia 241:925-929.17. WOODWARDw C. B., and R. F. A. Z",\ .s 197L The IN tc behav ior oif pure phuispholipase A,
and C toisards isitiuitiCallyr swollen erN thrcc tes and resealed ghoists. Bioc/himi. Riccplv.t. c274:272-278.
18. PARtWE. J. D.. and J. A. SPiisu-ta. 1982. Purification cit muscle actin. ic Methocds in CellBiology. Vol. 24. L. Wilson, editor. Academic Press. Nems Ycirk. 271-2S9.
19. Hi-MoCK. C. S.. and J. R. BANIM tR6. 1983. The quantitaticin cit G- and F-actiii in cultured
cells. Anal. Bioc 'lc',. 135:22-36.21). Z.SR. Jl. H. 1984. Bicistatistical Analysis. Prentice-Hall. Englesocid Clfs. Nem, Jerse\.21. Rciti.oiisi N .and H. J. SCII..v/NlvsN. 1977. The lipid requirement cifthe tCa ± Mg -
ATPase in the humnan erythrcct iieiibranc. asstudies bs saricus highly uiidpopolipases. hoc /io. Biccpiis . A, Id 464:17-36.
22. /w.SM. F. A., B. Rot wiin Ni. P. Comi itis. and 1.. L. M. V \\ Di i Nt \. 1975. Org~aniia-tion of phosphitlipids in human red cell membranes as detected by the acticon cit sariou s puritiedphosphcilipases. Biccc/imn. Biccphv , .c1ci 406:83 -96.
23. Wit LANOt. T. . and H. Fms I sric,. 1978. Anialcixins. phalicciciins. phallcisii. andi intdiiiaiite : The biiiloicallIy act ive coimpoinents oif" poiisonoius Aniaf mush roociii. C-. R. C-. Citicacl
Re'v. Bic'chern. 5-.185-260).24. Ccnt ((tc. L. M. . and L. G. Tii Nt N. 1984. Phallccidiii enhances actin assemibly by prekenting
nionomer dissociation. J. Cc/11 Riol. 99:529-535.

Actin and Myosin PLA2 Regulation 29
25. EstF..s, J. E., L. A. SF.tI-N. and L. C. GERSHMAN. 1981. Mechanism of action of phalloidinon the polymerization of muscle actin. Biochemistry 20:708-712.
26. CONRICOtE. K. M., and R. S. OcHs. 1987. Prwein interactions with phospholipase A. Fed.Proc. 46:2043.
27. POLLR.). T. D. Cytoplasmic contractile proteins. 1981. J. Cell Biol. 91:156s- 165s.
28. WILLINGHAM, M. C., K. M. YAMADA, S. S. YAMADA, J. P(O)YSSTF(;tR. and 1. PAS AN. 1977.Microfilament bundles and cell shape are related to adhesiveness to substratum and are dis-sociable from growth control in fibroblasts. Cell 10:37s-380.


















