
7. A NEW BIOSTRATIGRAPHIC INTERPRETATION OF THE ... · A NEW BIOSTRATIGRAPHIC INTERPRETATION OF THE...
9
7. A NEW BIOSTRATIGRAPHIC INTERPRETATION OF THE SEDIMENTARY RECORD RECOVERED AT SITE 462, LEG 61, NAURU BASIN, WESTERN EQUATORIAL PACIFIC 1 Isabella Premoli Silva, Dipartimento di Scienze della Terra, Universita di Milano 2 ABSTRACT Reinterpretation of the biostratigraphic record from Hole 462, drilled in the Nauru Basin during Leg 61, emphasizes the importance of larger foraminifers in dating heavily redeposited sequences. Younger ages derived from larger fora- minifers are supported by the occurrence of some planktonic marker species (foraminifers and radiolarians) previously interpreted as downhole contaminants. A major implication of the new younger ages is that the main erosional events previously dated at 37 Ma, 32 Ma, 26 Ma, and 13 Ma now have an age of 30 Ma, 28 Ma, 26 Ma or younger, and 12 Ma, respectively. Erosional events at the end of the Eocene and within the early Oligocene can no longer be documented. INTRODUCTION Two holes were drilled at Site 462 in the Nauru Basin during Leg 61 in 1978 by the Glomar Challenger. Hole 462A was reentered four years later in 1982 during Leg 89 in order to reach deeper horizons within the basaltic complex. The aim of this chapter is to reinterpret the biostratigraphic record from Hole 462, which was strong- ly biased by severe reworking and displacements of al- most all the age-diagnostic fossil groups (Larson, Schlang- er, et al., 1981), in light of a synthesis of the available data from both holes. DISCUSSION The sedimentary sequence recovered at Site 462 (Holes 462 and 462A) overlying the basaltic complex is charac- terized by pelagic clay and more rarely by radiolarian ooze, the autochthonous sediments, which alternate with car- bonate-rich layers. Pelagic clay is dominant in the lower portion of the sequence from 550 to 450 m sub-bottom (Cores 462-59 to 462-48), then it decreases upwards. In the Cenozoic, from 370 m to the top of the hole (Cores 462-39 to 462-1), the amount of pelagic clay and/or ra- diolarian ooze is about 10% of the total thickness. All the fossiliferous components, except the deep-dwell- ing agglutinated benthic foraminifers and possibly some radiolarians, have been transported. As a result, chro- nostratigraphy was jeopardized by reworking (Premoli Silva and Sliter, 1981; Premoli Silva and Violanti, 1981; Sanfilippo et al., 1981; Thierstein and Manivit, 1981). In the absence of age-diagnostic fossils from the autoch- thonous layers, the sediments were dated on the basis of the youngest components occurring in the various rese- dimented layers. However, under such conditions it is possible that even the youngest elements of a given fau- na and/or flora could be not only displaced but also re- Moberly, R., Schlanger, S. O., et al., Init. Repts. DSDP, 89: Washington (U.S. Govt. Printing Office). 2 Address: Dipartimento di Scienze della Terra, Universita di Milano, Via Mangiagalli, 34, 20133, Milan, Italy. worked. Consequently, the resulting dates could be much older than the actual age of deposition. Ages reported in the Site 462 report for Hole 462 sam- ples and in several other chapters as well (Larson, Sch- langer, et al., 1981) were based mainly on planktonic foraminifers and calcareous nannofossils. Discrepancies in the ages inferred from the different fossil groups are striking (see tables 4 and 5 in Larson, Schlanger, et al., 1981, pp. 52-53). Further discrepancies are evident if one compares the age inferred from the planktonics to those ages suggested by the shallow-water larger fora- minifers (Premoli Silva and Brusa, 1981). Most of the larger foraminifers are known to possess a high level of biostratigraphic resolution. Evolutionary lineages have been recognized within numerous, rapidly evolving genera. Moreover, each step within a lineage is identified as a species. Lineages are well dated by de- tailed quantitative studies and calibrated against strato- types (i. e., the Lepidorbitoides lineage). In some cases, their occurrence could be correlated with the planktonic foraminiferal zonation schemes (van Gorsel, 1978; Ad- ams, 1967; Schaub, 1951, 1981; Brönnimann, 1954; Haak and Postuma, 1975). Most of the species have a short stratigraphic range, and are diagnostic of specific time intervals. Furthermore, correctly identified specimens of a given evolutionary stage are age-diagnostic, even when recovered from a noncontinuous sequence. Thus the ages inferred from most of the larger foraminifers are as reli- able as those based on planktonic foraminifers, once the calibration of a lineage is established. At Site 462, although the shallow-water larger fora- minifers are displaced in deeper water as are the calcare- ous plankton and most of the radiolarians, the taxa found give younger minimum ages than did previous age as- signments that used planktonic organisms. The younger ages, based on the occurrence of some larger foramini- fers from Hole 462, are as follows (from bottom to top) (see Fig. 1): • Cores 462-52 and 462-51 contain numerous speci- mens of Vaughanina cubensis, which is Maestrichtian species according to Brönnimann (1954). Following Brön- nimann^ interpretation, Pseudorbitoides israelskyi among 311
-
Upload
truongkhanh -
Category
Documents
-
view
226 -
download
0
Transcript of 7. A NEW BIOSTRATIGRAPHIC INTERPRETATION OF THE ... · A NEW BIOSTRATIGRAPHIC INTERPRETATION OF THE...

7. A NEW BIOSTRATIGRAPHIC INTERPRETATION OF THE SEDIMENTARY RECORDRECOVERED AT SITE 462, LEG 61, NAURU BASIN, WESTERN EQUATORIAL PACIFIC1
Isabella Premoli Silva, Dipartimento di Scienze della Terra, Universita di Milano2
ABSTRACT
Reinterpretation of the biostratigraphic record from Hole 462, drilled in the Nauru Basin during Leg 61, emphasizesthe importance of larger foraminifers in dating heavily redeposited sequences. Younger ages derived from larger fora-minifers are supported by the occurrence of some planktonic marker species (foraminifers and radiolarians) previouslyinterpreted as downhole contaminants. A major implication of the new younger ages is that the main erosional eventspreviously dated at 37 Ma, 32 Ma, 26 Ma, and 13 Ma now have an age of 30 Ma, 28 Ma, 26 Ma or younger, and 12 Ma,respectively. Erosional events at the end of the Eocene and within the early Oligocene can no longer be documented.
INTRODUCTION
Two holes were drilled at Site 462 in the Nauru Basinduring Leg 61 in 1978 by the Glomar Challenger. Hole462A was reentered four years later in 1982 during Leg89 in order to reach deeper horizons within the basalticcomplex. The aim of this chapter is to reinterpret thebiostratigraphic record from Hole 462, which was strong-ly biased by severe reworking and displacements of al-most all the age-diagnostic fossil groups (Larson, Schlang-er, et al., 1981), in light of a synthesis of the availabledata from both holes.
DISCUSSION
The sedimentary sequence recovered at Site 462 (Holes462 and 462A) overlying the basaltic complex is charac-terized by pelagic clay and more rarely by radiolarian ooze,the autochthonous sediments, which alternate with car-bonate-rich layers. Pelagic clay is dominant in the lowerportion of the sequence from 550 to 450 m sub-bottom(Cores 462-59 to 462-48), then it decreases upwards. Inthe Cenozoic, from 370 m to the top of the hole (Cores462-39 to 462-1), the amount of pelagic clay and/or ra-diolarian ooze is about 10% of the total thickness.
All the fossiliferous components, except the deep-dwell-ing agglutinated benthic foraminifers and possibly someradiolarians, have been transported. As a result, chro-nostratigraphy was jeopardized by reworking (PremoliSilva and Sliter, 1981; Premoli Silva and Violanti, 1981;Sanfilippo et al., 1981; Thierstein and Manivit, 1981).In the absence of age-diagnostic fossils from the autoch-thonous layers, the sediments were dated on the basis ofthe youngest components occurring in the various rese-dimented layers. However, under such conditions it ispossible that even the youngest elements of a given fau-na and/or flora could be not only displaced but also re-
Moberly, R., Schlanger, S. O., et al., Init. Repts. DSDP, 89: Washington (U.S. Govt.Printing Office).
2 Address: Dipartimento di Scienze della Terra, Universita di Milano, Via Mangiagalli,34, 20133, Milan, Italy.
worked. Consequently, the resulting dates could be mucholder than the actual age of deposition.
Ages reported in the Site 462 report for Hole 462 sam-ples and in several other chapters as well (Larson, Sch-langer, et al., 1981) were based mainly on planktonicforaminifers and calcareous nannofossils. Discrepanciesin the ages inferred from the different fossil groups arestriking (see tables 4 and 5 in Larson, Schlanger, et al.,1981, pp. 52-53). Further discrepancies are evident ifone compares the age inferred from the planktonics tothose ages suggested by the shallow-water larger fora-minifers (Premoli Silva and Brusa, 1981).
Most of the larger foraminifers are known to possessa high level of biostratigraphic resolution. Evolutionarylineages have been recognized within numerous, rapidlyevolving genera. Moreover, each step within a lineage isidentified as a species. Lineages are well dated by de-tailed quantitative studies and calibrated against strato-types (i. e., the Lepidorbitoides lineage). In some cases,their occurrence could be correlated with the planktonicforaminiferal zonation schemes (van Gorsel, 1978; Ad-ams, 1967; Schaub, 1951, 1981; Brönnimann, 1954; Haakand Postuma, 1975). Most of the species have a shortstratigraphic range, and are diagnostic of specific timeintervals. Furthermore, correctly identified specimens ofa given evolutionary stage are age-diagnostic, even whenrecovered from a noncontinuous sequence. Thus the agesinferred from most of the larger foraminifers are as reli-able as those based on planktonic foraminifers, once thecalibration of a lineage is established.
At Site 462, although the shallow-water larger fora-minifers are displaced in deeper water as are the calcare-ous plankton and most of the radiolarians, the taxa foundgive younger minimum ages than did previous age as-signments that used planktonic organisms. The youngerages, based on the occurrence of some larger foramini-fers from Hole 462, are as follows (from bottom to top)(see Fig. 1):
• Cores 462-52 and 462-51 contain numerous speci-mens of Vaughanina cubensis, which is Maestrichtianspecies according to Brönnimann (1954). Following Brön-nimann^ interpretation, Pseudorbitoides israelskyi among
311

Age-diagnostic appearances
Spongaster tetras (R)
Artostrobium doliolum (R)
Globigerina nepenthes (P)
Hastigerina siphoniphera (P)
Hastigerina praesiphoniphera (P).
Stichocorys delmontensis (R)
2 0 0 -
Transported and reworked material
Reef skeletaldebris and larger
foraminifers
Rare shallow-water debris, miliolids,
Amphistegina, echmoid fragments. Age
Coeval ?
Miogypsinoides, Discocyclina,
Heterostegina, miliolids, bryozoan
debris. Age: Eocene, Oligocene.
Heterostegina borneensis, Nephrolepidina
sumatrensis, Eulepidina sp. cf.
E. ephippioides. Age: late Oligocene.
Nummulites pernotus, N. partschi, N.
burdigalensis minor. Assilina leymeriei.
Age: early Eocene. Nummulites
variolarius, N. bagelensis, Spiroclypeus
albapustula, S. vermicularis, Polylepidina
antillea, Asterocyclina matanzensis, A.
penuria, Miogypsinoides ubaghsi. Age:
late(?) Eocene. Nummulites vascus,
N. bouillei. Age: early Oligocene.
Pseudorbitoides. Age: late Campanian.
Rudistid fragments. Age:
mid-Cretaceous!?).
Few larger foraminifers, as in Cores 2 1 ,
22, and 32.
Planktonic andbathyal to abyssal
benthic foraminifers
Late Cretaceous through Pliocene trans-
ported planktonic foraminifers. Late
Miocene (N17) assemblages dominate.
Late Cretaceous through late Miocene
transported planktonic foraminifers.
Middle Miocene ( N 1 2 - N 1 3 ) assemblages
dominate.
Late Cretaceous, late Paleocene—early
Eocene, late Oligocene through early
Miocene transported planktonic
foraminifers. Early Miocene (N4, N7) and
middle Miocene ( N 1 0 - N 1 3 ) assemblages
dominate.
Late Cretaceous, late Paleocene—early
Eocene, late Oligocene, and early Miocene
(N7) transported planktonic foraminifers
with few small benthic foraminifers.
Late Oligocene and early Miocene
assemblages dominate.
Late Cretaceous, late Paleoceπe-early
Eocene, mid- and late Oligocene
transported planktonic foraminifers.
Mid-Oligocene assemblages dominate.
Late Cretaceous, late Paleocene—early
Eocene, early Oligocene transported
planktonic foraminifers wi th few small
benthic foraminifers. Early Oligocene
assemblages dominate.
Rare Aptian—Albian transportedplanktonic forminifers associated wi thLate Cretaceous, late Paleocene—earlyEocene, late Eocene displaced planktonicforaminifers.
Volcanicmaterial
Ash and volcanic glass.
Common volcanic glass and rock
fragments.
Abundant volcanic glass and rock
fragments.
Volcanic glass and rock fragments.
Age,Leg 61
s=5 .2
1
Age,this chapter
Pliocene oryounger
£ Oligocene

297
300
Globigerinita juvenilis (P)
Miogypsinoides ubaghsi (B) —fc•
Lepidorbitoides social'is (B)
Vaughanina cubensis (B)
A
* 2
Nummulites vascus, N. bouillei,
Miogypsinoides grandipustula. Age:
early Oligocene. Polylepidina sp.
aff. E. paucispira, Nummulites
problematicus, Asterocyclina
matanzeπsis, Miogypsinoides ubaghsi,
Spiroclypeus vermicularis, S. higginsi.
Age: latel?) Eocene. Assilina leymeriei,
Nummulites pernotus, N. burdigalensis
minor. Age: early Eocene. Orbitolinid
and rudistid fragments. Age:
mid-Cretaceous.
Asterocyclina matanzensis, several
Nummulites, Polylepidina sp. Age:
latel?) Eocene.
Assilina leymeriei, Nummulites pernotus.
Age: early Eocene.
Pseudorbitoides, Vaughanina? Age:
late Campanian.
Lepidorbitoides minor, L. socialis,
Orbitocyclina minima, Asterorbis
havanensis, A. rooki, A. cubensis,
Sulcoperculina vermunti, S. cubensis.
Age: middle Maestrichtian.
Lepidorbitoides bisambergensis. Age:
mid-early Maestrichtian. Pseudorbitoides
sp., Vaughanina sp. Age: late Campanian.
Pseudorbitoides israe/skyi, Vaughanina
cubensis, V. jordanae. Age: late
Campanian.
Late Cretaceous, late Paleocene, early
Eocene transported planktonic
foraminifers.
Late Cretaceous and late Paleocene (P4)
transported planktonic foraminifers.
Sporadic transported planktonic and
small bathyal benthic foraminifers.
First occurrence of transported planktonic
and small bathyal benthic foraminifers of
Cenomanian age.
Abundant volcanic glass and rock
fragments.
Very abundant volcanic glass, pyroxene
and rock fragments.
Abundant volcanic material: fresh to
altered glass, pyroxene, biotite, zeolite,
and rock fragments.
Abundant volcanic material: fresh to
altered glass, pyroxene, biotite, zeolite,
heavy opaque minerals, and very
abundant large rock fragments.
Paleo-cene
α> c
α
O
f i
Oligocene
Eocene
Campanian
e. Santon.Coniacian
Nannofossil silt toforaminifer sand
_ ^ _ Radiolarian sandI— *̂» \ and pelagic clay
Coarse layer
Chalk
— • -—| Mudstone and siltstone
Basalt
Figure 1. Distribution in DSDP Hole 462, Leg 61, of transported and reworked material after Premoli Silva and Brusa (1981) plotted against the new Leg 89 age interpretations,complemented by some age-diagnostic fossil appearances (in the first column, P = planktonic foraminifers, B = larger benthic foraminifers, and R = radiolarians).

I. PREMOLI SILVA
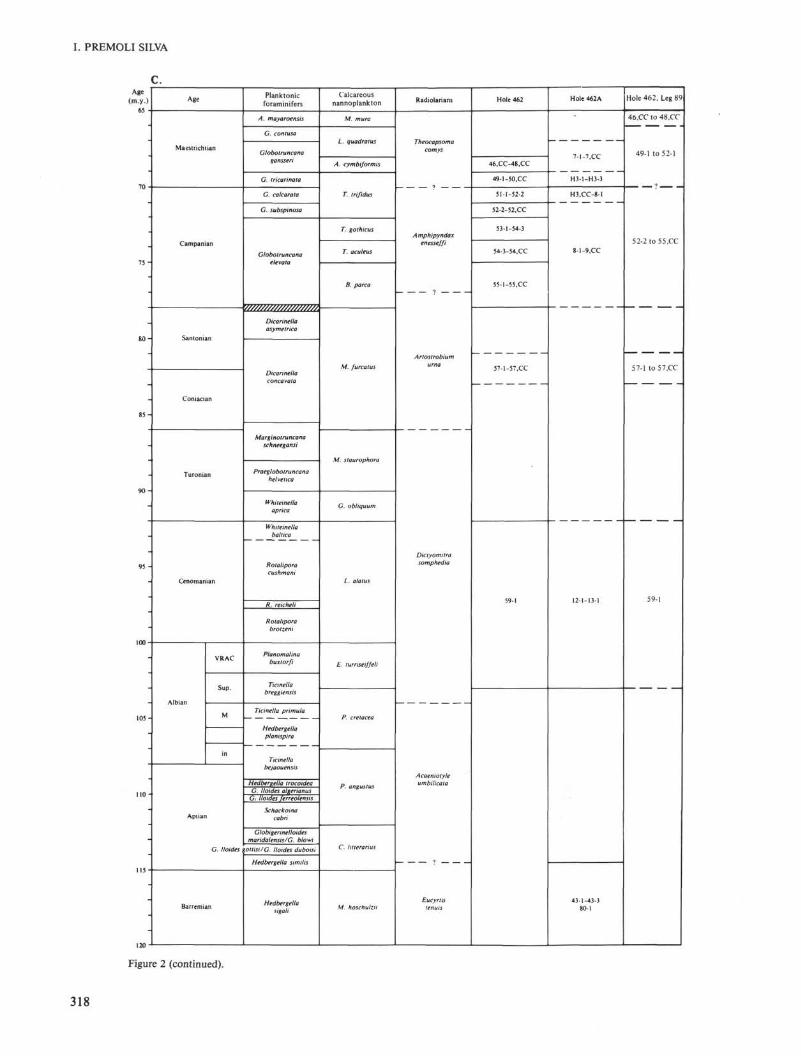
the larger foraminifers, and the planktonic foraminifersattributed to the Globotruncana subspinosa and Globo-truncana calcarata Zones, all diagnostic for the late Cam-panian, are reworked. The above-mentioned cores are thusdated as Maestrichtian and not late Campanian, as pre-viously reported (Larson, Schlanger, et al., 1981).
• Core 462-48 must be dated as late Maestrichtianbased on the occurrence of Lepidorbitoides socialis, theend member of the Lepidorbitoides lineage (van Gorsel,1978), and not as middle Maestrichtian, as inferred fromthe planktonic foraminifers attributed to the Globotrun-cana gansseri Zone. It is worth mentioning that the bio-stratigraphic signal from the calcareous nannofossilsspeaks in favor of an even older age (Thierstein andManivit, 1981).
• Core 462-34 contains a few specimens positively iden-tified as Miogypsinoides ubaghsi (Premoli Silva and Bru-sa, 1981). This genus is known to appear in the late Oli-gocene (Zone P21a = Globorotalia opima opimá). Thusall the planktonic components that are diagnostic of thelate Eocene are reworked.
• Section 462-32-1, again, contains representatives ofMiogypsinoides. It must be dated at least to Zone P21aof the late Oligocene as was Core 462-34, or even youn-ger. The planktonic foraminiferal assemblage of ZoneP20 (= Globigerina ampliapertura Zone) is consideredreworked.
On the basis of this age reinterpretation, some plank-tonic foraminifers that had been interpreted by PremoliSilva and Violanti (1981) as downhole contaminants mayin fact represent a correct record. The previously inter-preted contaminants that have now been reinstated asmarkers are (from bottom to top, see Fig. 1):
• Globorotalia opima opima, G. siakensis, well de-veloped globoquadrinids, Globigerina ciperoensis, andlarge Globoquadrina tripartita occur in several samplesfrom Core 462-34. Their presence is consistent with thelate Oligocene of Core 462-34 inferred from the occur-rence of Miogypsinoides. Moreover, some specimens ofGloborotalia kugleri also occur in the same samples: ifthese forms are also proved to belong to the host fauna,then Core 462-34 should be dated as latest Oligocene.Because of its abundance in the upper part of the se-quence, G. kugleri is still interpreted as a downhole con-taminant.
• Some specimens of Globigerinita juvenilis occur inSample 462-32-1, 73-75 cm. In the Atlantic this speciesappears within the range of Globigerina angulisuturalis(Zones P21b through P22) (author's personal observa-tion, 1984; Bolli, 1957). This occurrence would confirmnot only the late Oligocene age based on the presence ofMiogypsinoides (see above), but would also indicate thatCore 462-32 is younger than Core 462-34, as expected.
• Rare specimens of Hastigerina praesiphoniphera arerecorded in Sample 462-18-6, 25-27 cm. According toBlow (1969), this species appears within the early Mio-cene Zone N7. Consequently, the Oligocene/Mioceneboundary does not fall within Core 462-17, as stated inthe Site 462 report (Larson, Schlanger, et al., 1981) andin Premoli Silva and Violanti (1981).
• The subsequently recorded occurrences of Hasti-gerina siphoniphera in Sample 462-17-1, 6-8 cm, of Strep-tochilus pristinum in Sample 462-16-5, 40-41 cm, ofSphaeroidinellopsis subdehiscens, of evolutionary formstransitional to Globigerina nepenthes in Sample 462-15,CC, and finally of true Globigerina nepenthes in Sam-ple 462-14-4, 86-87 cm allow us to state that the middleMiocene interval from the top of Zone N12 to Zone N14is fully represented by Section 462-17-1 through Core462-14. In particular, Cores 462-17 and 462-16 are youn-ger than the early Miocene previously reported.
Besides the abundant reworking of the radiolarian fau-nas, Sanfilippo et al. (1981, p. 495) reported anomalousfirst appearances of some radiolarian species within theCenozoic sequence from Hole 462; in particular, theseauthors stated:
(1) The first appearance of Podocyrtis ampla amplaand P. ampla fasciolata is abnormally late in the Ceno-zoic sequence.
(2) The first appearance of Thyrsocyrtis bromia andCalocyclas turris is abnormally early in relation to thefirst appearance of Theocampe pirum, the Lithocycliaaristotelis group, Thyrsocyrtis tetracantha, and Carpo-canistrum azyx.
(3) The first appearance of Stichocorys delmontensisand Artostrobium doliolum is abnormally early.
(4) The isolated, abnormally early appearance of Sipho-campe corbula, Dictyocoryne ontongensis, and Spongas-ter tetras in Sample 462-15,CC can be explained only bydownhole contamination.
In view of the new foraminiferal data some anoma-lous occurrences can be reinterpreted as follows (see Rie-del and Sanfilippo, 1978) (Fig. 1):
• S. delmontensis from Core 462-20-4, 25-27 cm maybelong to the autochthonous assemblage that results inan early Miocene date for Core 462-20. The Oligocene/Miocene boundary, then, would be placed between Cores462-20 and 462-21.
• The occurrence of A. doliolum in Sample 462-14-2,86-88 cm is in agreement with the middle Miocene ageinferred for this core from planktonic foraminifers.
• Finally, if occurrence of S. tetras in Sample 462-15,CC is the result of downhole contamination, then theappearance of S. tetras in Sample 462-6-3, 65-67 cm isin agreement with the Pliocene age (or younger) inferredfrom the appearance of Globorotalia margaritae in Core462-7, the index species for the early Pliocene (PremoliSilva and Violanti, 1981).
The other anomalous occurrences mentioned by San-filippo et al. (1981) may be related to reworking similarto that of planktonic foraminifers.
Figure 2 shows the new age interpretations plottedagainst those reported in Larson, Schlanger, et al. (1981).It appears that ages derived from calcareous nannofos-sils are consistently older than those based on plankton-ic foraminifers and radiolarians.
The reinterpretation of the planktonic foraminiferalfaunas emphasizes the importance of larger foramini-fers as biostratigraphic indicators. In particular, at Hole462 they are more reliable than planktonic foraminifers
314
NEW BIOSTRATIGRAPHIC INTERPRETATION OF SEDIMENTARY RECORD
in dating heavily redeposited sequences. A possible ex-planation is that the age-diagnostic planktonic faunasare too diluted by reworked faunas to be perceived dur-ing a routine biostratigraphic observation. In contrast,the shallow-water components, being displaced sudden-ly by exceptional events, were not significantly contami-nated.
The main consequences of the younger ages now re-ported from the interval from Cores 462-34 to 462-14are that (1) the accumulation rates plotted in table 1 byPremoli Silva and Violanti (1981) must be recalculatedas reported in Table 1, and (2) the erosional events previ-ously dated at 37 Ma, 32 Ma, 26 Ma, and 13 Ma nowhave an age of 30 Ma, 28 Ma, 26 Ma or younger, and 12Ma, respectively. The erosional event at the end of theEocene and within the early Oligocene can no longer bedocumented.
REFERENCES
Adams, C. G., 1967. Tertiary foraminifera in the Tethyan, American,and Indo-Pacific Provinces. System. Ass. Publ., 7:195.
Blow, W. H., 1969. Late middle Eocene to Recent planktonic forami-niferal biostratigraphy. Proc. First Int. Conf. Planktonic Microfos-sils, Geneva, 1967, 1:199-422.
Bolli, H. M., 1957. Planktonic Foraminifera from the Oligocene-Mio-cene Cipero and Lengua formations of Trinidad, B. W. I. U.S.Nat. Museum Bull., 215:97-124.
Brönnimann, P., 1954. Upper Cretaceous orbitoidal foraminifera fromCuba. II. Vaughanina Palmer. Contrib. Cush. Found. Foraminif.Res., 5:91.
Haak, R., and Postuma, J. A., 1975. The relation between the tropicalplanktonic foraminiferal zonation and the Tertiary Far East letterclassification. Geol. en Mijnbouw, 54(34): 195.
Larson, R. L., Schlanger, S. O., et al., 1981. Init. Repts. DSDP, 61:Washington (U.S. Govt. Printing Office).
Premoli Silva, I., and Brusa, C , 1981. Shallow-water skeletal debrisand larger foraminifers from Deep Sea Drilling Project Site 462,Nauru Basin, Western Equatorial Pacific. In Larson, R. L., Schlang-er, S. O., et al., Init. Repts. DSDP, 61: Washington (U.S. Govt.Printing Office), 439-473.
Premoli Silva, I., and Sliter, W. V., 1981. Cretaceous planktonic fora-minifers from the Nauru Basin, Leg 61, Site 462, Western Equato-rial Pacific. In Larson, R. L., Schlanger, S. O., et al., Init. Repts.DSDP, 61: Washington (U.S. Govt. Printing Office), 423-437.
Premoli Silva, I., and Violanti, D., 1981. Cenozoic planktonic fora-minifer biostratigraphy of Deep Sea Drilling Project Hole 462, Na-uru Basin (Western Equatorial Pacific), and distribution of the pe-lagic components. In Larson, R. L., Schlanger, S. O., et al., Init.Repts. DSDP, 61: Washington (U.S. Govt. Printing Office), 397-422.
Riedel, W. R., and Sanfilippo, A., 1978. Stratigraphy and evolutionof tropical Cenozoic radiolarians. Micropaleontology, 23(l):61-97.
Sanfilippo, A., Westberg, M. J., and Riedel, W. R., 1981. Cenozoicradiolarians at Site 462, Deep Sea Drilling Project Leg 61, WesternTropical Pacific. In Larson, R. L., Schlanger, S. O., et al., Init.Repts. DSDP, 61: Washington (U.S. Govt. Printing Office), 495-505.
Schaub, H., 1951. Stratigraphic and Palaeontologie des Schlieren-flysches. Schweiz. Palaeont. Abh., 68:108.
, 1981, Nummulites et Assilines de la Téthys paléogène. Tax-inomie, phylogenése et biostratigraphic Schweiz. Palaeont. Abh.,104:1-238, and atlas.
Thierstein, H. R., and Manivit, H., 1981. Calcareous-nannofossil bio-stratigraphy, Nauru Basin, Deep Sea Drilling Project Site 462, andUpper Cretaceous nannofacies. In Larson, R. L., Schlanger, S. O.,et al., Init. Repts. DSDP, 61: Washington (U.S. Govt. Printing Of-fice), 475-494.
van Gorsel, J. J., 1978. Late Cretaceous orbitoidal foraminifera. InHedley, R. H., and Adams, C. G. (Eds.), Foraminifera (Vol. 3):London (Academic Press), 1-120.
Date of Initial Receipt: 16 November 1984Date of Acceptance: 26 April 1985
Table 1. Recalculated accumulation rates of the Cenozoic sequence from Hole 462, Leg 61, according to the newage interpretations reported in Figures 1 and 2.
Core-section Age
ClayDuration Thickness thickness
(m.y.) (m) (m)
Totalaccumulation rate
(m/m.y.)
Transported materialaccumulation rate
(m/m.y.)
1 to 3 Pleistocene 1.8 28.5 10.65 15.83 24.28
4 to 8
10 to 12-512-6 to 14-414-5 to 16-516,CC to 17-117-2 to 18.CC19 to 20
21 to 23-223-3 to 32-132-2 to 34.CC35 to 38.CC
Miocene
Oligocene
Pliocene?
N16/N17N14NI3 (top)N12 (top)N7N5
P22P21bP21aP20/P18?
2.2
3.00.80.50.31.01.5
1.02.02.07.0
38.0
26.517.511.03.5
17.019.0
22.084.027.038.0
14.15
2.360.080.101.783.155.38
0.250.17
4.83
17.30
8.8421.8722.0011.7017.0012.67
22.0042.0013.505.43
30.38
8.7422.0022.2414.1020.2214.16
22.3042.2713.5015.28
315

I. PREMOLI SILVA
A.Age
(m.y.)0
early
early
early
Planktonic foraminifers
Oligocene
Core 4-Core 8
Core 10, Section 1
Core 12, Section 5
Core 12, Section 6-Core 14.CC. Core 1A
-Nil NN6-
Core 16-Core 17, Section 1
Core 17, Section 3-Core 18
Calcareous nannofossils
Core 1, Sections 1-6
Core 1,CC
Core 5.CC
Core 10-Core 12,Section 1 Core 1A
Core 12-Core 15
Core 16, Section 1
Core 16,CC-Core 18
Lamprocyrti•
haysi
Pterocaniumprismatiufn
Spongasiepen tas
Snchocorysperegnna
Ommatartuspenulumus
Ommalarlusantepenulnmus
Cannartuspetlerssoni
Dorcadospynsalara
Calocycletlacoslala
Slichocoryswolffii
Snchocorysdelmonleπsis
Cyrtocapsellatetrapera
Core I-Core 3,CC
Core 4, Section 2-Core 5, Section 1
Core 5, Section 2-Core 6, Section 5
Core 7, Section I-Core 8, Section 4
Core I A, Section I-Core 1A.CC
Core 8.CC-Core 12, Section 4
Core 15, Section 1-Core 15.CC
Core 16, Section 2
Core 17, Section I-Core 17, Section 6
Core 18, Section 2
Hole 462, Leg 89
19-1 to 20.CC
Figure 2. A-C. Neogene through Cretaceous biostratigraphy of Hole 462, Leg 61. Note the new age interpretations (last column)compared with those reported in Larson, Schlanger, et al. (1981). Hachured areas = uncertain boundaries.
316

NEW BIOSTRATIGRAPHIC INTERPRETATION OF SEDIMENTARY RECORD
Age(m.y.)
25 -
Age Planktonic foraminifers Calcareous nannofossils RadiolariansHole 462.
Leg 89
26 -
27 -
28 -
29 -
3 0 -
35 -
40 -
45 -
50 -
55 -
Oligocene
Eocene
Paleocene
60 -
65
C. ciperoensisP22
late
'Gr.' opima opimaP21 "~
G. ampliaperturaP20
early
Cassigerinellachipolensis/
pseudohastigerinamicro
P19-P18
T. cerroazulensis
lateGlobigerinatheka
semiinvoluta
Truncorotaloidesrohri
Orbulinoidesbeckmanni
middle Morozovellalehneri
Globigerinathekasubconglobata
Hantkeninaaragonensis
Acarininapentacamerata
early
Morozovellaaragonensis
M. formosa
M. subbotinae
M. edgan
Morozovellavelascoensis
late Ptanorotalitespseudomenardii
M. pusilla pusilla
M. angulata
M. uncinata
early "M. " trinidadensis
Subbotinapseudobulloides
"P. " eugubina
Core 19-Core 23,Section 2
Core 23, Section 3-Core 27, Section 2
Core 27, Section 3-Core 32, Section 3
Core 32, Section 4-Core 33
Core 34
NP25
NP24
NP23
NP22
NP21
NP20
NP19
NP18
NP17
NP16
NP15
NP14
NP13
NP12
NP10
NP9
NP8
NP7
NP6
NP5
NP4
NP3
NP2
NP1
Core 19, Section 2-Core 19, Section 6
D elongaia
Core 19,CC-Core 22
Dorcadospyrisateuchus
Core 23-Core 32, Section 2
Core 2A
Core 32,CC-Core 33
Core 34-Core 38
Theocyrtisluberosa
Thyrsocyrtisbromia
Podocyrlisgoetheana
Podocyrtis chalara
Core 39, Section 1Podocyrtis
mitra
Podocyrtis ampla
Core 39, Section 3-Core 39.CC
Core 41-Core 42
Thyrsocyrtistriacantha
T. mongolfieriT. cryptocephala
Phormocyrtisstriata
Buryellaclinata
Core 44, SectionCore 5A.CC
Core 45,CC
Core 18, Section 6
Core 19, Section 2-Core 25.CC
Core 2A, Section 1-Core 2A.CC
Core 27, Section 2-Core 37.CC
Core 38, Section I-Core 38.CC
Core 39, Section 2-Core 39, Section 5
Core 41.CC
21-1 to23-2
23-3 to32-1
32-2 to34,CC
35-1 to38.CC (?)
39-1 to40.CC (?)
41.CC
42-1 and43.CC (?)
44-1 to44,CC
45.CC
Figure 2 (continued).
317
I. PREMOLI SILVA
C.Age
(m.y.)
65
AgePlanktonic
foraminifers
Calcareousnannoplankton
Hole 462, Leg 89
Campanian
A. mayaroensis
Clobolruncanagansseri
A. cymbiformis
T. irifidus
C. subspin
T. gothi<
Clobolruncanaelevala
B. parca
Dicarinellaasymetrica
Dicarinellacoπcavala
M. furcatus
Marginotrunschneegan
M. slauropho
Praeglobotrunchelvetica
Wnttemellaaprica
C. obliquun
Whiieinellabaltica
Rotaliporacushmani
Roialiporabrolzem
buxtorfi E. lumseif/eli
Sup. Tianellabreggiensis
Ticinella primula
Hedbergellaplanispira
Ticinella
Hedbergella irocoideaG. lloides algerianus
P. aπgustus
i lloides ferreolensis
Globigerinelloides
mandalensis/G. blow
oltisi/G. lloides duboisi
Hedbergella similis
Hedbergellasigali
46,CC to 48,CC
Theocapsomacomys
46.CC-48.CC7-1-7,CC
H3-1-H3-3
H3.CC-8-1
Amphipyndaxenessef/i 52-2 toS5,CC
57-1 to57,CC
Diclyomilr,somphedia
Acaeniolyleumbilkaia
Eucyrttslenuis
43-1-43-380-1
Figure 2 (continued).
318

NEW BIOSTRATIGRAPHIC INTERPRETATION OF SEDIMENTARY RECORD
Plate 1. 1-3, 9. Miogypsinoides ubaghsi Tan. Sample 462-32-1, 5-10 cm, late Oligocene, (1) tangential section, × 75, (2) equatorial section, mar-ginal portion, × 62, (3) equatorial section, × 60, (9) equatorial section, × 55. 4-6. Vaughanina cubensis Palmer. Sample 462-52-1, 98-101 cm,Maestrichtian, (4) equatorial section, × 50, (5-6) fragments of the marginal portion, external view, × 60. 7-8. Lepidorbitoides socialis (Ley-merie). Sample 462-48 soup, late Maestrichtian, (7) detail of the marginal portion, equatorial section, × 190, (8) equatorial section, × 95.
319


















