
4147 (1): 059 066 Article ZOOTAXA · species (Uetz & Hallermn 2016). Members of the genus are...
8
Accepted by C. Siler: 23 Jun. 2016; published: 2 Aug. 2016 ZOOTAXA ISSN 1175-5326 (print edition) ISSN 1175-5334 (online edition) Copyright © 2016 Magnolia Press Zootaxa 4147 (1): 059–066 http://www.mapress.com/j/zt/ Article 59 http://doi.org/10.11646/zootaxa.4147.1.3 http://zoobank.org/urn:lsid:zoobank.org:pub:0FB6E658-DEAA-47D7-89BF-9C07C650FEC7 On the taxonomy and phylogeny of the skinks Lipinia sekayuensis Grismer, Ismail, Awang, Rizal, & Ahmad and Lipinia surda Boulenger from Peninsular Malaysia L. LEE GRISMER 1 , PERRY L. WOOD JR. 2 , MUHAMMAD F. SYAFIQ 3 , BAIZUL H. BADLI-SHAM 3 , SYED A. RIZAL 3 , AMIRRUDIN B. AHMAD 3 & EVAN S. H. QUAH 4 1 Department of Biology, La Sierra University, 4500 Riverwalk Parkway, Riverside, California, 92515-8247 USA. E-mail: [email protected] 2 Department of Biology, Brigham Young University, 150 East Bulldog Boulevard, Provo, Utah 84602 USA. E-mail: [email protected] 3 School of Marine and Environmental Sciences, Universiti Malaysia Terengganu, 21030 Kuala Terengganu, Terengganu, Malaysia. E-mail: [email protected] 4 School of Biological Sciences, Universiti Sains Malaysia, 11800 USM, Pulau Pinang, Penang, Malaysia. E-mail: [email protected] Abstract An integrative taxonomic analysis based on additional specimens and color photographs of Lipinia sekayuensis and addi- tional color photographs of L. surda from Pulau Tioman and the Gunung Panti Forest Reserve, Peninsular Malaysia con- firm the previous hypotheses that L. sekayuensis is a valid species and is the sister species of L. surda. The two species share a 12.8% sequence divergence between them. Key words: Hutan Lipur Sekayu, Lipinia,Scincidae, Malaysia, phylogeny, taxonomy Introduction The genus Lipinia Boulenger is a modest radiation of Southeast Asian and Oceanian skinks that contains at least 28 species (Uetz & Hallermn 2016). Members of the genus are distributed from the Andaman and Nicobar Islands of India, eastward through Thailand and Cambodia to the Philippines, and southward through the Thai-Malay Peninsula and Singapore to Sumatra and Borneo (Linkem 2013). This extensive distribution continues across many islands of the Indo-Australian Archipelago to New Guinea (Günther 2000), and onward throughout much of the South Pacific (Adler et al. 1995). Many species of Lipinia are generally small, diurnal, somewhat secretive, terrestrial skinks that spend time foraging through surface debris and in low shrubs searching for small arthropods (Grismer, 2011a). Others, however, are arboreal, rainforest species with strikingly bold color patterns and are commonly seen moving up and down the trunks of large trees, often several meters above the ground. Recently, the genus has been shown to be a widely polyphyletic assemblage within the sphenomorphine tribe Sphenomorphini (Linkem 2013), and until a stable phylogenetic taxonomy is established for the tribe, the generic classification of the contents of Lipinia will remain unsettled. Grismer et al. (2014) described Lipinia sekayuensis Grismer, Ismail, Awang, Rizal, & Ahmad from Hutan Lipur Sekayu, Terengganu, Peninsular Malaysia on the basis of a single specimen and in the absence of molecular and color pattern data from living material. They hypothesized that L. sekayuensis likely is most closely related to L. surda Boulenger based on the fact that both species lack an external ear opening and have a dull, nearly unicolor color pattern. They indicated that the two species differed from one another in the number of paravertebral scales; ventral scales; suubdigital lamellae on Toes III–V; size of the transverse subcaudal scales; and subtle differences in the dorsal striping pattern. We have since acquired two additional specimens of L. sekayuensis: an adult (SVL=38 mm) from Sungai Bubu (AHH 466) near the type locality at Hutan Lipur Sekayu and a juvenile (SVL=25 mm)
Transcript of 4147 (1): 059 066 Article ZOOTAXA · species (Uetz & Hallermn 2016). Members of the genus are...

ZOOTAXA
ISSN 1175-5326 (print edition)
ISSN 1175-5334 (online edition)Copyright © 2016 Magnolia Press
Zootaxa 4147 (1): 059–066
http://www.mapress.com/j/zt/Article
http://doi.org/10.11646/zootaxa.4147.1.3
http://zoobank.org/urn:lsid:zoobank.org:pub:0FB6E658-DEAA-47D7-89BF-9C07C650FEC7
On the taxonomy and phylogeny of the skinks Lipinia sekayuensis Grismer,
Ismail, Awang, Rizal, & Ahmad and Lipinia surda Boulenger from Peninsular
Malaysia
L. LEE GRISMER1, PERRY L. WOOD JR.2, MUHAMMAD F. SYAFIQ3, BAIZUL H. BADLI-SHAM3,
SYED A. RIZAL3, AMIRRUDIN B. AHMAD3 & EVAN S. H. QUAH4 1Department of Biology, La Sierra University, 4500 Riverwalk Parkway, Riverside, California, 92515-8247 USA.
E-mail: [email protected] of Biology, Brigham Young University, 150 East Bulldog Boulevard, Provo, Utah 84602 USA. E-mail: [email protected] of Marine and Environmental Sciences, Universiti Malaysia Terengganu, 21030 Kuala Terengganu, Terengganu, Malaysia.
E-mail: [email protected] of Biological Sciences, Universiti Sains Malaysia, 11800 USM, Pulau Pinang, Penang, Malaysia.
E-mail: [email protected]
Abstract
An integrative taxonomic analysis based on additional specimens and color photographs of Lipinia sekayuensis and addi-
tional color photographs of L. surda from Pulau Tioman and the Gunung Panti Forest Reserve, Peninsular Malaysia con-
firm the previous hypotheses that L. sekayuensis is a valid species and is the sister species of L. surda. The two species
share a 12.8% sequence divergence between them.
Key words: Hutan Lipur Sekayu, Lipinia,Scincidae, Malaysia, phylogeny, taxonomy
Introduction
The genus Lipinia Boulenger is a modest radiation of Southeast Asian and Oceanian skinks that contains at least 28
species (Uetz & Hallermn 2016). Members of the genus are distributed from the Andaman and Nicobar Islands of
India, eastward through Thailand and Cambodia to the Philippines, and southward through the Thai-Malay
Peninsula and Singapore to Sumatra and Borneo (Linkem 2013). This extensive distribution continues across many
islands of the Indo-Australian Archipelago to New Guinea (Günther 2000), and onward throughout much of the
South Pacific (Adler et al. 1995). Many species of Lipinia are generally small, diurnal, somewhat secretive,
terrestrial skinks that spend time foraging through surface debris and in low shrubs searching for small arthropods
(Grismer, 2011a). Others, however, are arboreal, rainforest species with strikingly bold color patterns and are
commonly seen moving up and down the trunks of large trees, often several meters above the ground. Recently, the
genus has been shown to be a widely polyphyletic assemblage within the sphenomorphine tribe Sphenomorphini
(Linkem 2013), and until a stable phylogenetic taxonomy is established for the tribe, the generic classification of
the contents of Lipinia will remain unsettled.
Grismer et al. (2014) described Lipinia sekayuensis Grismer, Ismail, Awang, Rizal, & Ahmad from Hutan
Lipur Sekayu, Terengganu, Peninsular Malaysia on the basis of a single specimen and in the absence of molecular
and color pattern data from living material. They hypothesized that L. sekayuensis likely is most closely related to
L. surda Boulenger based on the fact that both species lack an external ear opening and have a dull, nearly unicolor
color pattern. They indicated that the two species differed from one another in the number of paravertebral scales;
ventral scales; suubdigital lamellae on Toes III–V; size of the transverse subcaudal scales; and subtle differences in
the dorsal striping pattern. We have since acquired two additional specimens of L. sekayuensis: an adult (SVL=38
mm) from Sungai Bubu (AHH 466) near the type locality at Hutan Lipur Sekayu and a juvenile (SVL=25 mm)
Accepted by C. Siler: 23 Jun. 2016; published: 2 Aug. 2016 59

from the Sungai (=river) Pur, approximately 5.5 km to the north. Both specimens were accompanied with color
photographs and we sequenced both for mitochondrial gene ND2. Additionally, we have acquired color
photographs of living specimens of L. surda from Pulau Tioman, Pahang and the Gunung Panti Forest Reserve,
Johor (Fig. 1).
We want to be clear that the Lipinia phylogeny used herein does not fully represent the complete evolutionary
history of this poloyphyletic genus without including several other genera (Linkem, 2013). The phylogeny was
constructed from GenBank sequence data and from sequences generated from the only tissue samples available for
Lipinia and was used solely to test the hypotheses of species validity of L. sekayuensis and its relationship to L.
surda (Table 1).
FIGURE 1. Upper left: Principal Component Analysis of Lipinia sekayuensis and L. surda. Right: Maximum clade credibility
topology from Bayesian analyses showing the relationships among the two focal species of Lipinia and a suite of other selected
taxa based on recent higher-level analyses (Linkem 2013). Nodal support values are Bayesian posterior probabilities/Maximum
Likelihood bootstrap values. Lower left: Distribution of Lipinia sekayuensis and L. surda in Peninsular Malaysia and the east
coast islands of Tioman and Aur in the Seribuat Archipelago.
Materials and methods
Phylogenetic analysis. A dataset was composed of 1,035 bp of the mitochondrial gene ND2 and constructed from
21 individuals comprising 11 species of Lipinia that formed the ingroup and two individuals of Papuascincus
stanleyanus as outgroup taxa based on Linkem et al. (2011; Table 1). Total genomic DNA was isolated from liver
or skeletal muscle from specimens stored in 95% ethanol using the Qiagen DNeasyTM tissue kit (Valencia, CA,
USA). Only the mitochondrial gene NADH dehydrogenase subunit 2 (ND2) was amplified using a double-stranded
Polymerase Chain Reaction (PCR) under the following conditions: 1.0 ml genomic DNA (~10–33 mg), 1.0 ml (10
mM) light strand primer (5’-AAGCAGTTGGGCCCATACC-3’: Macey et al. 1997), 1.0 µl (10 mM) heavy strand
primer (5’-AACCAAACCCAACTACGAAAAAT-3’; Macey et al. 1997), 1.0 ml deoxynucleotide triphosphate
(DNTPs) pairs (1.5 mM), 2.0 ml 5x buffer (1.5 mM), 1.0 ml MgCl 10x buffer (1.5 mM), 0.18 ml Taq polymerase
GRISMER ET AL.60 · Zootaxa 4147 (1) © 2016 Magnolia Press

(5u/ml), and 7.5 ml H2O. PCR reactions were run on an Eppendorf Mastercycler gradient thermocycler under the
following conditions: initial denaturation at 95°C for 2 min, followed by a second denaturation at 95°C for 35 s,
annealing at 52°C for 35 s, followed by a cycle extension at 72°C for 35 s, for 33 cycles. All PCR products were
visualized via electrophoresis on a 1% agarose gel. Successfully amplified PCR products were vacuum purified
using MANU 30 PCR Millipore plates and purified products were resuspended in sterile molecular grade water.
Purified PCR products were sequenced using the ABI Big-Dye Terminator v3.1 Cycle Sequencing Kit in an ABI
GeneAmp PCR 9700 thermal cycler. Cycle sequencing reactions were purified with Sephadex G-50 Fine (GE
Healthcare) and sequenced on an ABI 3730xl DNA Analyzer at the BYU DNA Sequencing Center. All sequences
were edited in Geneious v6.1.8 (Drummond et al. 2011) and aligned by eye. Mesquite v3.0.2 (Maddison &
Maddison 2015) was used to check for premature stop codons and to ensure the correct amino acid read frame.
The phylogenetic analysis applied two model-based methods, Maximum Likelihood (ML) and Bayesian
Inference (BI). The Bayesian Information Criterion (BIC) implemented in IQ-TREE (Nguyen et al. 2015)
calculated TVM+I+G4, TPM3u+G4, and K3Pu+G4 to be the best-fit models of evolution for the first, second, and
third codon positions, respectively. Maximum Likelihood analyses using IQ-TREE employed 1,000 bootstrap
pseudoreplicates via the ultrafast bootstrap approximation algorithm. A partitioned Bayesian analysis was carried
out in MrBayes v3.2.6 (Huelsenbeck & Ronquist 2001; Ronquist et al. 2012) on CIPRES (Miller et al. 2010) using
default priors. Models of evolution used were approximated from those calculated for the ML analysis and thus
employed GTR+I+G for all codon positions. Two simultaneous Markov Chain Monte Carlo (MCMC) runs were
performed with four chains per run (three hot and one cold) using default priors. The analysis was run for two
million generations, sampled every 200 generations, and halted after the average standard deviation split frequency
was below 0.01 as indicated on the output. The first 25% of the trees were discarded as burnin using the sumt
function. Nodes having ultrafast ML bootstrap values of 90 and BI posterior probabilities of 0.95 and above were
considered strongly supported (Huelsenbeck et al. 2001; Nguyen et al. 2015; Wilcox et al. 2002).
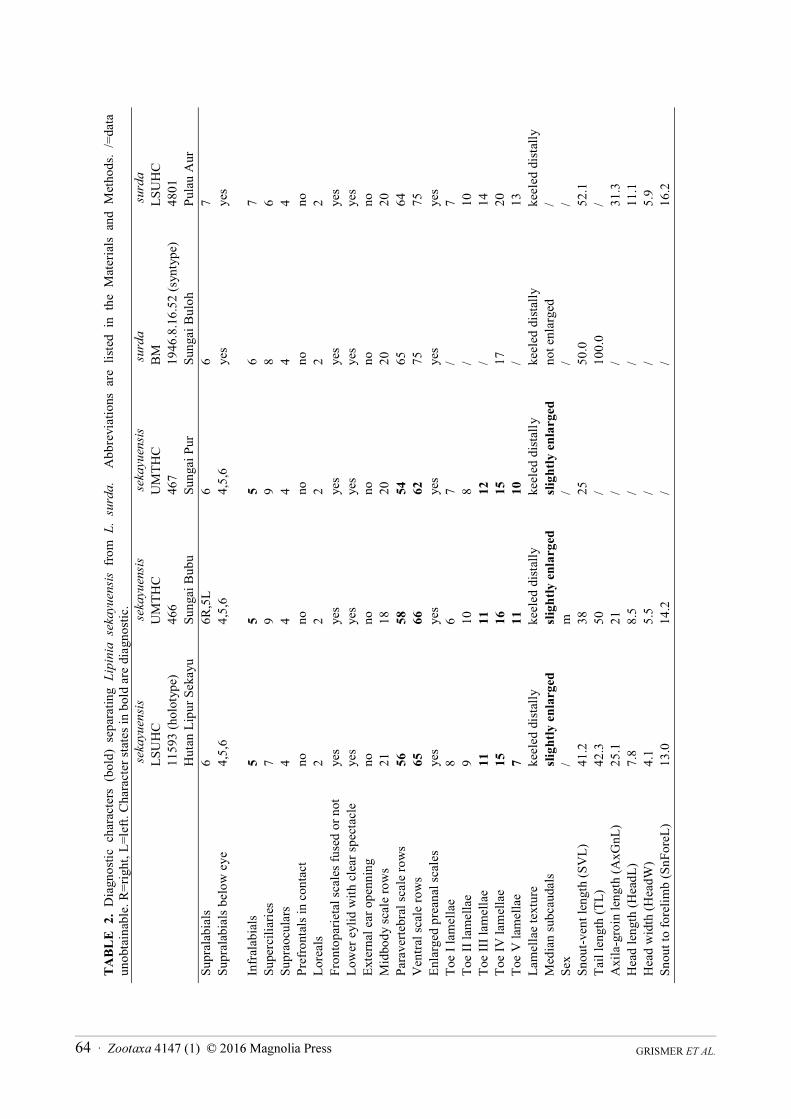
Morphological analysis. Scale terminology follows Grismer et al. (2009, 2014) and Lim (1998). All
measurements were made by LLG to the nearest 0.1 mm using Mitutoyo digital calipers. Scale counts were made
on the right side of the body with a Nikkon SMZ 1500 dissecting microscope. Measurements and scale counts
taken were snout–vent length (SVL) measured from the tip of the rostral scale to the vent; tail length (TailL)
measured from the tip of the tail (original or regenerated) to the vent; axilla–groin length (AxGnL) measured from
the posterior margin of the forelimb insertion to the anterior margin of the hind limb insertion; head length (HeadL)
measured from the anterior margin of the ear opening to the tip of the rostral scale; head width (HeadW) measured
as the widest portion of the temporal region; snout to forelimb length (SnForeL) measured from the anterior margin
of the forelimb insertion to the tip of the rostral scale; midbody scale rows counted as the number of longitudinal
scale rows encircling the body at a point midway between the limb insertions; paravertebral scale rows counted as
the number of scales in a line from the parietal scales to a point on the dorsum opposite the vent; and ventral scale
rows counted as a row of scales between the postmentals and the cloacal plate. Other standard counts include
supraoculars, suboculars, loreals, supralabials, infralabials, and lamellae beneath the toes. Additional characters
examined were the degree of contact between the prefrontal scales, state of fusion between frontoparietal scales,
presence or absence of enlarged precloacal scales, and texture of subdigital lamellae. Color pattern characters
examined were the numbers of stripes on the flanks and dorsum and the degree of darkness of those stripes.
A principle component analysis (PCA) was implemented by the prcomp command in R v 3.2.1 (R Core Team
2015) to find the overall best low-dimensional representation of the significant morphological variation in the data.
Scale count characters used in the PCA were suprlabials, infralabials, superciliaries, midbody scales, paravertebral
scales, ventral scales, and fourth toe lamellae (Table 2). In order to normalize the distribution of all characters, the
data were log-transformed prior to analysis.
Abbreviations for institutions are FNMH—Field Museum of Natural History, Chicago, Illinois, USA; JAM—
Jimmy A. McGuire, University of California, Berkeley, Berkeley, California, USA; KU—University of Kansas,
Lawrence, Kansas, USA; RMB—Rafe M. Brown, University of Kansas, Lawrence, Kansas, USA; CAS—
California Academy of Sciences, San Francisco, California, USA; MVZ—Museum of Vertebrate Zoology,
University of California, Berkeley, Berkeley, California, USA; LSUHC—La Sierra University Herpetological
Collection, La Sierra University, Riverside, California, USA; TNHC—Texas Natural History Collection,
University of Texas at Austin, Austin, Texas; ) UMTHC—Universiti Malaysia Terengganu Herpetological
Collection, Universiti Malaysia Terengganu, Kuala Terengganu, Malaysia, and LSUDPC—La Sierra University
Digital Photo Collection, La Sierra University, Riverside, California, USA.
Zootaxa 4147 (1) © 2016 Magnolia Press · 61LIPINIA SEKAYUENSIS & LIPINIA SURDA

����
���������������� ����������������������������
���
����
������
�
���
��
� �
� �
�������
���
��
Papu
asci
ncus
stan
leya
nus
�����������
�������!�"����#�$���%�����#�&���'�(�����#�&���'�(����%��
)*+������
Papu
asci
ncus
stan
leya
nus�
�����������
�������!�"����,�$���%�����,�&���'�(�����#�&���'�(����%��
)*+������
� ����
���
��
Lipi
nia
noct
ua
�����-�+.+�
�����/���#��%�����/���#��%��������(����%��
)*+����0�
Lipi
nia
pulc
hella
1�2��.�-���
��������#�3�����/���#�3����� ��4���#�$
������������ �3����#�3���%���5�4������
)*+������
Lipi
nia
pulc
hella
1�2��.�-���
��������#�3�����/���#�3����� ��4���#�$
������������ �3����#�3���%���5�4������
)*+������
Lipi
nia
vitti
gera
*$�2���0��.
1�������
&6+0+����
Lipi
nia
vitti
gera
7�82�������
9���$
������#� ��%#� ����� ��%�
&6+0+����
Lipi
nia
vitti
gera
7�82�����0�
���:����#� ����� ��4����
&6+0+��-�
Lipi
nia
surd
a 7�82��+�0��
9���$
������#�)����#� ��������
&6+0+��+�
Lipi
nia
seka
yuen
sis
��2�+���
9���$
������#�1���%%��#���%���3�:��
&6+0+��.�
Lipi
nia
seka
yuen
sis
��2�+���
9���$
������#�1���%%��#���%��� ���
&6+0+����
Lipi
nia
relic
ta
$(;��-�-���
�������#�&����
����<%%��#� �����<%%��#�4�������� �4����%��� �$���'���
&6+0+����
Lipi
nia
infr
alin
eola
ta
)�$���--�
����!��#�&��������7����#�&�:������&����'��8����#� ��4�������!���1�%%����
&6+0+����
Lipi
nia
infr
alin
eola
ta
)�$������
����!��#�&��������3���#�&�:������3�%%��#� ��4�������!���1�%���
&6+0+����
Lipi
nia
inex
pect
ata
7�82�����-�
<���$
������#���:��#�1������/����
&6+0+��0�
Lipi
nia
inex
pect
ata
7�82�����+�
<���$
������#���:��#�1������/����
&6+0+����
Lipi
nia
quad
rivi
rgat
a 5$3�-�0��
��������=�$������ �/�#�$������/���#���4�������� ��4���#�$������������ ������
&6+0+����
Lipi
nia
quad
rivi
rgat
a 5$3�-�0��
��������#�(����� �/�#���%���/���#���%���>������� ��4���#�$������������ �(������
&6+0+��-�
Lipi
nia
quad
rivi
rgat
a &8�-0���0�
��������#�(����� �/�#���:��/���#���:�� ��4���#�$������������ �������
&6+0+��+�
�
GRISMER ET AL.62 · Zootaxa 4147 (1) © 2016 Magnolia Press

Results
The ML and BI analyses recovered trees with the exact same topology supporting Lipinia sekayuensis and L. surda
as sister species with 1.00/100 (BI/ML) nodal support (Fig. 1). Additionally, we observe a 12.8% uncorrected
pairwise sequence divergence for ND2 between samples of the two focal taxa. The morphological analysis of the
additional specimens of L. sekayuensis continue to support its discrete distinction from L. surda in having five as
opposed to six or seven infralabials; 54–58 as opposed to 64 or 65 paravertebrals; 62–66 as opposed to 75 ventrals;
11 or 12 as opposed to 14 subdigital lamellae on Toe III; 15 or 16 as opposed to 17–20 subdigital lamellae on Toe
IV; 7–10 as opposed to 13 subdigital lamellae on Toe V; and slightly enlarged, transverse, median subcaudals as
opposed to no enlarged, transverse subcaudals (Table 2).
Principle Component Analysis revealed wide separation between L. sekayuensis and L. surda along the first
principle component which accounted for 68% of the total variation and loaded most heavily for infralabials, Toe
IV lamellae, and ventral scales (Table 3). From the color photographs of the adult L. sekayuensis (UMTHC 466)
and L. surda from Pulau Tioman (LSUDPC 10063) and Pulau Aur (LSUDPC 6191), it is clear that adult L.
sekayuensis and L. surda cannot be differentiated on the basis of coloration and pigmentation pattern (Fig. 2). Both
species have an orange to red iris; a gold-brown dorsum; a series of small white spots on the labials and the sides of
the head, neck, and flanks; and the posterior portion of the tail is gray and covered with white flecks. The juvenile
L. sekayuensis (UMTHC 467) has a markedly bold color pattern similar to that of the adults with the exception of
the entire tail being black and covered with transversely oriented white spots forming rings. The differences in faint
dorsal striping between the two species reported by Grismer et al. (2012) are far too subtle and variable in living
material to be of diagnostic value.
FIGURE 2. Upper left: Adult Lipinia sekayuensis (UMTHC 466) from Sungai Bubu, Terengganu, Peninsular Malaysia;
photograph by S. Rizal. Middle left: Juvenile Lipinia sekayuensis (UMTHC 467) from Sungai Pur, Terengganu, Peninsular
Malaysia; photograph by S. Rizal. Lower Adult left: Lipinia surda (LSUHC 4801) from Pulau Aur, Johor, Peninsular Malaysia;
photograph by L. Grismer. Upper right: Adult Lipinia surda (LSUDPC 10063) from Pulau Tioman, Pahang, Peninsular
Malaysia; photograph by B. Schembri. Lower right: Juvenile Lipinia surda (LSUDPC 10064) from Gunung Panti Forest
Reserve, Johor, Peninsular Malaysia; photograph by C. Foley.
Zootaxa 4147 (1) © 2016 Magnolia Press · 63LIPINIA SEKAYUENSIS & LIPINIA SURDA
����
�������%���������������?:���@��������%�
Lipi
nia
seka
yuen
sis ����
L. s
urda
. �
::��4����������������������$�����������$�������AB�����
��:����:����5B��%��#�LB�� ��������������������:�����������%������
� se
kayu
ensi
s s
ekay
uens
is
seka
yuen
sis
surd
a s
urda
7�82��
8$12��
8$12��
3$�
7�82���
���.�-�?��������@�
+���
+���
��+�������.��?�����@�
+�0��
�2����7�������'����
��%���3�:��
��%��� ���
��%���3�����
���������
�������:����
��
�5#.7�
��
��
��
�������:����b
elow
eye
�
+#.#��
+#.#��
+#.#��
���
���
/ ����:����
����
����
��
��������������
��
��
��
��
��
������������
+�
+�
+�
+�
+�
�� ���������������
��
��
��
��
��
7������
��
��
��
��
��
*������������������ ����������
���
���
���
���
���
7�!���������!�������������������
���
���
���
���
���
<C��������������%��
��
��
��
��
��
$��:�����������!�
���
���
�0�
�0�
�0�
���4����:�����������!�
���
���
� �
�.�
�+�
(�������������!�
���
���
���
�.�
�.�
<���%���������������
���
���
���
���
���
1���/����������
��
��
��
A���
1���//����������
��
�0�
��
A��0�
1���///����������
���
���
���
A��+�
1���/(����������
���
���
���
���
�0�
1���(����
������
!���
��"
�A�
�-�
7����������C�����
'��������������
'��������������
'��������������
'��������������
'��������������
$������:�������
#$%��
�$&�� $���'�
#$%���$&�� $���'�
#$%��
�$&�� $���'�
�������%���
A�
��C�
A���
A�A�
A�
����D4�����%���?�(7@�
+����
-��
�.�
.0�0�
.����
1������%���?17@�
+��-�
.0�
A��00�0�
A�
�C���D%������%���?�C"7@�
�.���
���
A�A�
-��-�
2������%���?2���7@�
����
��.�
A�A�
�����
2����!�����?2���9@�
+���
.�.�
A�A�
.���
�������� ������:�?�*���7@�
�-�0�
�+���
A�A�
�����
�
GRISMER ET AL.64 · Zootaxa 4147 (1) © 2016 Magnolia Press

TABLE 3. Summary statistics and principal component scores for male and female morphological variables.
Discussion
The morphological, color pattern, and molecular analyses of the additional two specimens of Lipinia sekayuensis,
coupled with the additional color photographs of L. surda, enabled us to test and accept the hypotheses of the
separate specific identity and the sister relationship between these two species. Based on only a photograph of a
specimen from the Gunung Panti Forest Reserve, Johor, in southern Peninsular Malaysia, we cannot unequivocally
place it in either species. However, being that L. surda occurs on both sides of the Banjaran (=mountain range)
Titiwangsa (Grismer 2011a; Grismer et al. 2014) at Sungai Buloh in Kuala Lumpur and the islands of Tioman and
Aur in the Seribuat Archipelago (Grismer 2011a,b), we tentatively consider this specimen as L. cf. surda (Fig. 1).
Many other lowland species of lizards endemic to Peninsular Malaysia have a similar distribution pattern (see
distribution maps in Grismer 2011a).
More importantly, this analysis underscores the long held precept that if the morphological data are clear
enough—i.e. character state distinction is sufficiently evident and the utility of those characters to delimit species
within that taxonomic group has been well-established—proposing a robust, testable hypothesis of specific identity
is advisable. In fact, it would require more scientific justification to not describe the population in question as a
new species. In many cases, rare or secretive species may not be collected for decades (or ever again) and to leave
these populations unnamed provides them little or no formal conservation protection. The alternative of
inadvertently describing a junior synonym is little more than an annoying issue of bookkeeping—not so much an
issue of science. And it is far less problematic than not providing conservation measures to a species that could
become threatened or may already be threatened due to microendemism. This is especially true in megadiverse
biodiversity hotspots such as Southeast Asian rainforests where the rate of deforestation in most places is
increasing faster that our ability to catalog its diversity.
Acknowledgements
Department of Forestry Terengganu is gratefully acknowledged for granting us permission to conduct research at
Hutan Lipur Sekayu. MFS, BHBS, LHBI, MTA, SAR, and ABA would like to thank the School of Marine and
Environmental Sciences, Universiti Malaysia Terengganu for providing facility and financial support to conduct
the field survey. We thank Nurul Asyikin, Nurdiyanah and Nur Amirah for helping us during the field survey.
Genetic sequencing was supported in part by a NSF grant EF-1241885 issued to Jack W. Sites Jr.
References
Adler, G.H., Austin, C.C. & Dudley, R. (1995) Dispersal and speciation of skinks among archipelagos in the tropical Pacific
PC1 PC2 PC3 PC4 PC5
Standard deviation 2.184113216 1.279686341 0.619359762 0.456558664 1.78E-15
Proportion of variance 0.68148 0.23394 0.0548 0.02978 0
Cumulative proportion 0.68148 0.91542 0.97022 1 1
Eigenvalues 4.77035054 1.637597132 0.383606514 0.208445813 3.17E-30
Suprlabials 0.362570402 -0.428475623 -0.190287933 -0.529145292 -0.604471562
Infralabials 0.449073806 0.090712211 -0.122341131 -0.299994937 0.472991549
Superciliaries -0.377098875 0.302422989 0.351046073 -0.773122709 0.128460368
Midbody scale rows 0.139131921 -0.704827715 0.495353913 -0.001436932 0.438612941
Paravertebrals 0.392493532 0.328205046 0.476721969 0.086021202 -0.162615618
Ventrals 0.415445497 0.253174423 0.424039158 0.114232301 -0.221972423
Toe IV lamellae 0.422691417 0.219368004 -0.416102467 -0.108806448 0.355378883
Zootaxa 4147 (1) © 2016 Magnolia Press · 65LIPINIA SEKAYUENSIS & LIPINIA SURDA

Ocean. Evolutionary Ecology, 9, 529–541.
http://dx.doi.org/10.1007/BF01237834
Drummond, A.J., Ashton, B., Buxton, S., Cheung, M., Cooper, A., Duran, C., Field, M., Heled, J., Kearse, M., Markowitz, S.,
Moir, R., Stones-Havas, S., Sturrock, S., Thierer, T. & Wilson, A. (2011) Geneious V5.6, Available from: http://
www.Geneious.com/ (Accessed 1 Aug. 2016)
Grismer, L.L. (2011a) Lizards of Peninsular Malaysia, Singapore and Their Adjacent Archipelagos. Edition Chaimira,
Frankfürt am Main. 728 pp.
Grismer, L.L. (2011b) Amphibians and Reptiles of the Seribuat Archipelago (Peninsular Malaysia). Edition Chaimira,
Frankfürt am Main. 239 pp.
Grismer, L.L., Ismail, L.H.B, Awang, M.T., Rizal, S.A. & Ahmad, A.B. (2014) A new species of lowland skink (genus Lipinia
Gray, 1845) from northeastern Peninsular Malaysia. Zootaxa, 3821 (4), 457–464.
http://dx.doi.org/10.11646/zootaxa.3821.4.4
Grismer, L.L., Norhayai, A. & Chan, K.O. (2009b) A new, diminutive, upland Sphenomorphus Fitzinger 1843 (Squamata;
Scincidae) from the Belum-Temengor Forest Complex, Peninsular Malaysia. Zootaxa, 2312, 27–38.
Günther, R. (2000) In alten sammlungen aus Neuguinea entdeckt: zwei neue arten der gattung Lipinia (Squamata: Scincidae).
Salamandra, 36, 157–174.
Huelsenbeck, J.P. & Ronquist, F. (2011) MrBayes: Bayesian Inference of Phylogenetic Trees. Bioinformatics (Oxford), 17,
754–755.
Lim, L.J. (1998) The taxonomy of West Malaysian and Singapore Scincidae (Reptilia: Sauria). Unpublished M.S. Thesis,
National University of Singapore, Kent Ridge, Singapore, 234 pp.
Linkem, C.W., Diesmos, A.C. & Brown, R.M. (2011) Molecular systematics of the Philippine forest skinks (Squamata:
Scincidae: Sphenomorphus): testing morphological hypotheses of interspecific relationships. Zoological Journal of the
Linnean Society 163, 1217–124.
http://dx.doi.org/10.1111/j.1096-3642.2011.00747.x
Linkem, C.W. (2013) Molecular Phylogenetics and Biogeography of Sphenomorphini (Squamata: Scincidae). PhD Thesis.
Department of Ecology and Evolutionary Biology, University of Kansas. 250 pp.
Macey, J.R., Schulte, J.A., Larson, A., Tuniyev, B.S., Orlov, N. & Papenfuss, T.J. (1997) Molecular phylogenetics, tRNA
evolution, and historical biogeography in anguid lizards and related taxonomic families. Molecular phylogenetics and
evolution, 12 (3), 250–272.
http://dx.doi.org/10.1006/mpev.1999.0615
Maddison, W. & Maddison, D. (2015) Mesquite: a modular system for evolutionary analysis. version 3.04. 2015. Avaliable
from: http://mesquiteproject.org (Accessed 1 Aug. 2016)
Miller, M.A., Pfeiffer, W. & Schwartz, T. (2010) “Creating the CIPRES Science Gateway for inference of large phylogenetic
trees” in Proceedings of the Gateway Computing Enviroments Workshop (GCE), 14 Nov. 2010, New Orleans, LA, pp. 1–
8.
http://dx.doi.org/10.1109/GCE.2010.5676129
Nguyen, L.-T., Schmidt, H.A., von Haeseler, A. & Minh, B.Q. (2015) IQ-TREE: A fast and effective stochastic algorithm for
estimating maximum likelihood phylogenies. Molecular Biology and Evolution, 32, 268–274.
http://dx.doi.org/10.1093/molbev/msu300
R Core Team. (2015) R: A Language and Environment for Statistical Computing. Ver 3.2. R Foundation for Statistical
Computing. Vienna: Austria. Available from: www.R-project.org/ (Acessed 13 Jul. 2016)
Ronquist, F., Teslenko, M., van der Mark, P., Ayres, D.L., Darling, A., Höhna, B. Larget,B., Liu, L., Suchard, M.A. &
Huelsenbeck, J.P. (2012) Mr. Bayes 3.2: Efficient Bayesian Phylogenetic Inference and Model Choice across a Large
Model Space. Systematic Biology, 61, 539–542.
http://dx.doi.org/10.1093/sysbio/sys029
Uetz, P. & Hallermann, J. (2016) The Reptile Database. Avaliable from: http://www.reptile-database.org (Accessed 1 Aug.
2016)
Wilcox, T.P., Zwickl, D.J., Heath, T.A. & Hillis, D.M. ( 2002) Phylogenetic relationships of the Dwarf Boas and a comparison
of Bayesian and bootstrap measures of phylogenetic support. Molecular Phylogenetics and Evolution, 25, 361–371.
http://dx.doi.org/10.1016/S1055-7903(02)00244-0
GRISMER ET AL.66 · Zootaxa 4147 (1) © 2016 Magnolia Press


















![STEMfest panel Sept 2015[4147]](https://static.fdocuments.net/doc/165x107/58722a271a28ab3b7a8b5c37/stemfest-panel-sept-20154147.jpg)