
41 & 42 - Nucleic Acid Metabolism
56
NUCLEOTIDE METABOLISM LECTURE 41 &42
-
Upload
mada-claws -
Category
Documents
-
view
24 -
download
0
Transcript of 41 & 42 - Nucleic Acid Metabolism

NUCLEOTIDE METABOLISM
LECTURE 41 &42

Objectives
At the end of these lectures, students should be able
to:
• Describe synthesis of deoxyribonucleotides
• Compare purine and pyrimidine synthesis
• Compare purine and pyrimidine degradation

Both the salvage and de novo synthesis pathways of purine
and pyrimidine biosynthesis lead to production of
nucleoside-5'-phosphates through the utilization of an
activated sugar intermediate and a class of enzymes called
phosphoribosyltransferases.
The activated sugar used is 5-phosphoribosyl-1-
pyrophosphate, PRPP.
PRPP is generated by the action of PRPP synthetase and
requires energy in the form of ATP as shown:
Nucleotide Metabolism

Nucleotide Metabolism

There are two distinct pathways leading to purines:
1. The de novo pathway (from scratch)
2. Salvage pathways (take free base and create a nucleotide)
3. Conversion of ribonucleotides into deoxyribonucleotides
The major site of purine synthesis is in the liver.
Dietary absorption of ingested purines and pyrimidines is very
low (most of the purines are converted to uric acid in the
intestine)
Purine Synthesis

Origin of the atoms in the purine ring

Synthesis of the purine nucleotides begins with PRPP and
leads to the first fully formed nucleotide, inosine 5'-
monophosphate (IMP)
Purine Synthesis
Synthesis of Inosine Monophosphate
• Basic pathway for biosynthesis of purine ribonucleotides
• Starts from ribose-5-phosphate which is derived from the
pentose phosphate pathway
• Requires 11 steps overall
• occurs primarily in the liver

Steps 1 thru 3
• Step 1:Activation of ribose-5-phosphate – enzyme: PRPP synthetase
– product: 5-phosphoribosyl-a-pyrophosphate (PRPP)
– PRPP is also a precursor in the biosynthesis of pyrimidine
nucleotides and the amino acids histidine and tryptophan

Step 2: commited step
Acquisition of purine atom 9
enzyme: Gln: PRPP amido transferase
displacement of pyrophosphate group by glutamine amide nitrogen
product: b-5-phosphoribosylamine
Steps 1 and 2 are tightly regulated
by feedback inhibition

Steps 3
Acquisition of purine atoms C4,
C5, and N7
– enzyme: glycinamide
ribotide synthetase
– b-phosphoribosylamine
reacts with ATP and glycine
– product: glycinamide
ribotide (GAR)

Steps 4 thru 6
Step 4: acquisition of purine atom C8
– formylation of free a-amino group of GAR
– enzyme: GAR transformylase
– co-factor of enzyme is N10-formyl THF
Step 5: acquisition of purine atom N3
– The amide amino group of a second glutamine is transferred to form formylglycinamidine ribotide (FGAM)
Step 6:
• closing of the imidazole ring or formation of 5-aminoimidazole ribotide


Step 7
Acquisition of C6
– C6 is introduced as HCO3-
– enzyme: AIR carboxylase (aminoimidazole ribotide
carboxylase)
– product: CAIR (carboxyaminoimidazole ribotide)
– enzyme composed of 2 proteins: PurE and PurK (synergistic
proteins)

Steps 8
Acquisition of N1
– N1 is acquired from
aspartate in an amide
condensation reaction
– enzyme: SAICAR
synthetase
– product: 5-
aminoimidazole-4-(N-
succinylocarboxamide)ri
botide (SAICAR)
– reaction is driven by
hydrolysis of ATP
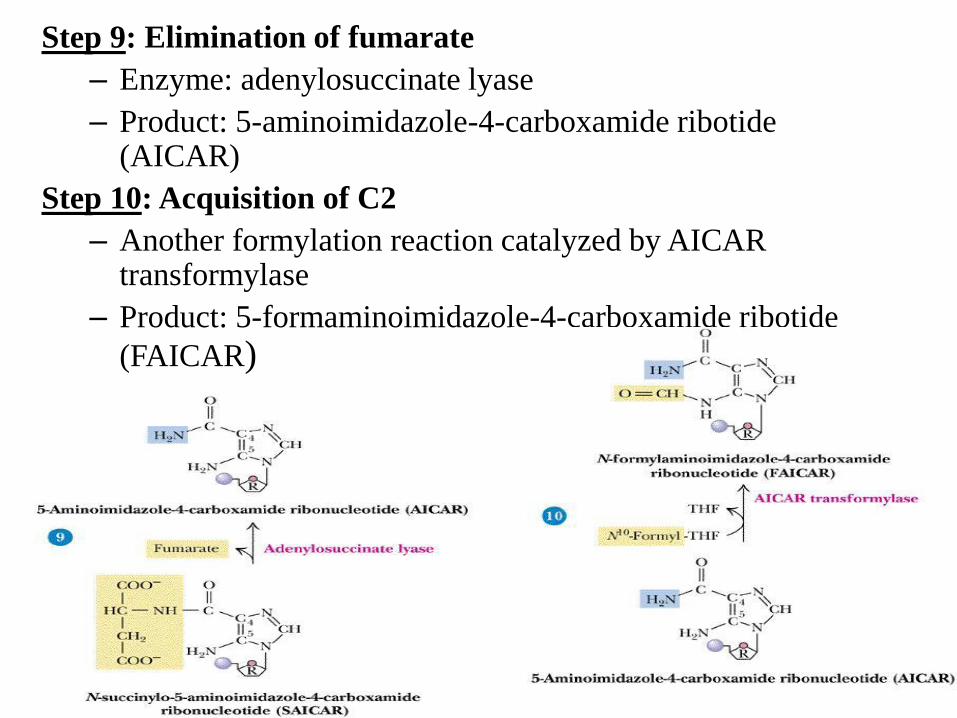
Step 9: Elimination of fumarate
– Enzyme: adenylosuccinate lyase
– Product: 5-aminoimidazole-4-carboxamide ribotide (AICAR)
Step 10: Acquisition of C2
– Another formylation reaction catalyzed by AICAR transformylase
– Product: 5-formaminoimidazole-4-carboxamide ribotide
(FAICAR)

Step 11
Cyclization or ring closure to form IMP
• water is eliminated
• in contrast to step 6 (closure of the imidazole ring), this reaction does not require ATP hydrolysis
• once formed, IMP is rapidly converted to AMP and GMP (it does not accumulate in cells


Outline of Purine
Ring Biosynthesis
10 steps are required:
Two ring closure

Biosynthesis of AMP from IMP
Notes
1- GTP is required for AMP synthesis
2- Need to add –NH2 group to
position six of IMP; use the succinate
to fumarate reaction (urea cycle)
3- Adenylosuccinate synthetase is
regulated by AMP levels

Biosynthesis of GMP from IMP

Conversion of AMP, GMP to more
Phosphorylated Form

Conversion of IMP to AMP and GMP

- to be incorporated into DNA and RNA, nucleoside
monophosphates (NMP’s) must be converted into
nucleoside triphosphates (NTP’s)
- nucleoside monophosphate kinases (adenylate & guanylate kinases)
- nucleoside diphosphate kinase
AMP + ATP 2 ADP
GMP + ATP GDP + ADP
accomplished by separate enzymes
GDP + ATP GTP + ADP
same enzyme acts on all nucleotide di & triphosphates
nucleoside diphosphate kinase is an enzyme which plays
a key role in the activation of antiviral nucleosides such as Retrovir/AZT
Purine nucleoside diphosphates and triphosphates

Regulatory Control of Purine
Nucleotide Biosynthesis • GTP is involved in AMP synthesis and ATP is involved in
GMP synthesis (reciprocal control of production)
• PRPP is a biosynthetically “central” molecule (why?)
– ADP/GDP levels – negative feedback on Ribose Phosphate
Pyrophosphokinase
– Amidophosphoribosyl transferase is activated by PRPP levels
– APRT activity has negative feedback at two sites
• ATP, ADP, AMP bound at one site
• GTP,GDP AND GMP bound at the other site
• Rate of AMP production increases with increasing
concentrations of GTP; rate of GMP production increases with
increasing concentrations of ATP

Regulatory Control of Purine Biosynthesis
• Above the level of IMP production:
– Independent control
– Synergistic control
– Feedforward activation by PRPP
• Below level of IMP production
– Reciprocal control
• Total amounts of purine nucleotides controlled
• Relative amounts of ATP, GTP controlled

The purine salvage pathway
• Purine bases created by degradation of RNA or DNA and intermediate of purine synthesis were costly for the cell to make, so there are pathways to recover these bases in the form of nucleotides
• Two phosphoribosyl transferases are involved:
– APRT (adenine phosphoribosyl transferase) for adenine
– HGPRT (hypoxanthine guanine phosphoribosyl transferase) for guanine or hypoxanthine

Salvage of purines
O
HH
OHOH
CH2
H O
OP
O
OH
OH
H
P
O
O-
O P
O
O-
O-
O
HH
OHOH
CH2
H H
OP
O
OH
OH N N
NN
NH2
adenine
PPi
Adenine
phosphoribosyltransferase
(APRT)

Salvage of purines
• Salvage is needed to maintain the purine pool (biosynthesis is
not completely adequate, especially in neural tissue)
• Hypoxanthine-guanine phosphoribosyltransferase (HGPRT)
• Hypoxanthine + PRPP IMP + Ppi
• Guanine + PRPP GMP + Ppi
• Lack of HGPRT leads to Lesch-Nyhan syndrome. Lack of
enzyme leads to overproduction of purines which are
metabolized to uric acid, which damages cells

Salvage of purine bases

Synthesis of Pyrimidine Ribonucleotides
• Shorter pathway than for purines
• Base is made first, then attached to ribose-P (unlike purine biosynthesis)
• Only 2 precursors (aspartate and glutamine, plus HCO3
-) contribute to the 6-membered ring
• Requires 6 steps (instead of 11 for purine)
• The product is UMP (uridine monophosphate)

Origin of the atoms in the pyrimidine ring
Pyrimidine Biosynthesis
Important differences from Purine Biosynthesis: 1- Pyrimidine base is syntheized before the ribose is added
2- PRPP, Gln, CO2, and aspartate are required for both purine and
pyrimidine biosynthesis
3- There is both de novoand salvage pathways for pyrimidine
synthesis, although the salvage pathway is not as important as it is
for purine recycling.

Step 1: Synthesis of carbamoyl phosphate
Condensation of glutamine, bicarbonate in the presence of ATP
Carbamoyl phosphate synthetase exists in 2 types: CPS-I which
is a mitochondrial enzyme and is dedicated to the urea cycle
and arginine biosynthesis) and CPS-II, a cytosolic enzyme
used here
CPS-II is the major site of regulation in animals: UDP and
UTP inhibit the enzyme and ATP and PRPP activate it
It is the committed step in animals

Step 2: Synthesis of Carbamoyl Aspartate
Enzyme is aspartate transcarbamoylase (ATCase),
catalyzes the condensation of carbamoyl phosphate with aspartate
with the release of Pi
ATCase is the major site of regulation in bacteria; it is activated by
ATP and inhibited by CTP
Carbamoyl phosphate is an “activated” compound, so no energy
input is needed at this step
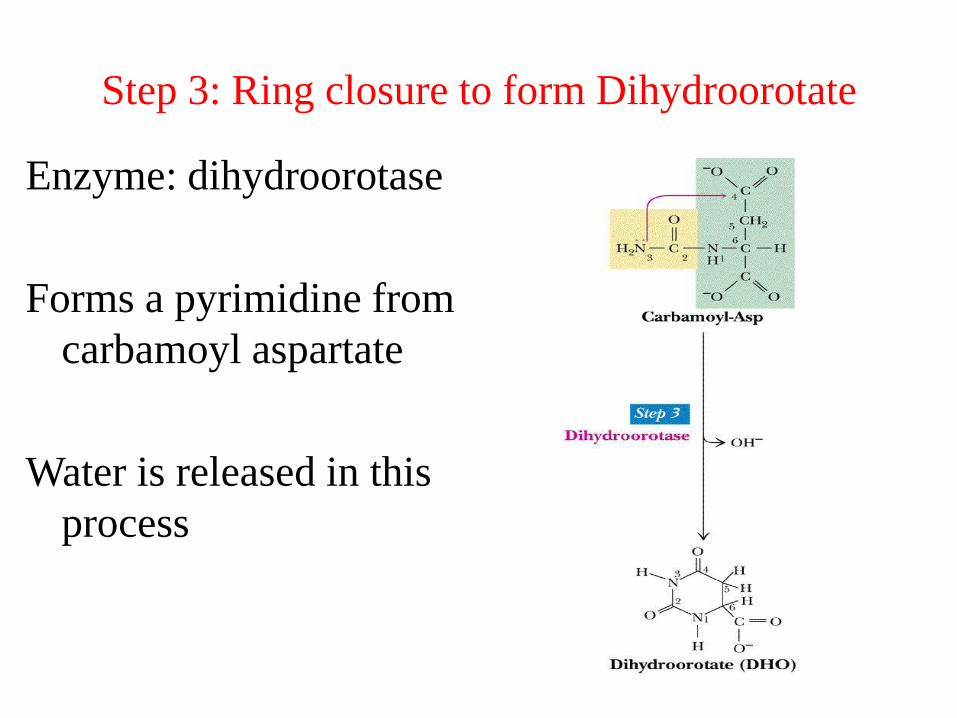
Step 3: Ring closure to form Dihydroorotate
Enzyme: dihydroorotase
Forms a pyrimidine from
carbamoyl aspartate
Water is released in this
process

The first 3 enzymatic reactions are catalyzed by 3 separate proteins/enzymes in E. coli
In animals, all 3 steps are found in a multifunctional enzyme (210 kD). This allows “channeling” of the substrates and products between active sites without releasing them to the medium where they could be degraded.
The acronym CAD is used as a name for the multienzyme: carbamoyl phosphate synthetase, aspartate transcarbamoylase and dihydroorotase
Channeling also increases the overall rate of multistep processes
Step 1-3: Pyrimidine Synthesis

Step 4: Oxidation of Dihydroorotate to Orotate
An irreversible reaction
Enzyme: dihydroorotate dehydrogenase
Oxidizing power is derived from quinones (thru
coenzyme Q)

Step 5: Acquisition of Ribose phosphate moiety
Enzyme: orotate phosphoribosyl transferase
Ribose phosphate originates from PRPP
Product is orotidine-5’-monophosphate (OMP)
Orotate phosphoribosyl transferase is also used in salvage of uracil and cytosine to their corresponding nucleotide
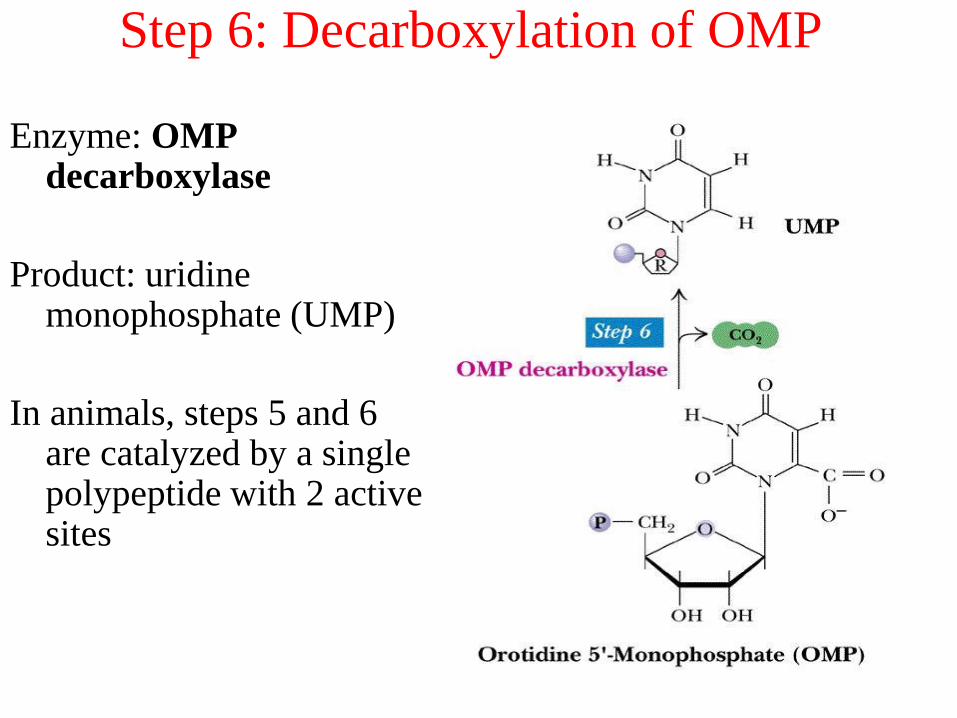
Step 6: Decarboxylation of OMP
Enzyme: OMP decarboxylase
Product: uridine monophosphate (UMP)
In animals, steps 5 and 6 are catalyzed by a single polypeptide with 2 active sites

Regulatory Control of Pyrimidine Synthesis
Differs between bacteria and animals
Bacteria – regulation at ATCase rxn
Animals – regulation at carbamoyl phosphate synthetase II - UDP and UTP inhibit enzyme; ATP and PRPP activate it
- UMP and CMP competitively inhibit OMP Decarboxylase
Purine synthesis inhibited by ADP and GDP at ribose phosphate pyrophosphokinase step, controlling level of PRPP also regulates pyrimidines

Regulation of pyrimidine nucleotide biosynthesis
Glutamine +
HCO3- +
ATP
Carbamoyl
phosphate
carbamoyl phosph.
synthetase
Orotate
OMPUMPUTP + CTP
orotate
phosphoribosyl
transferase
UTP and CTP are feeback inhibitors of CPS II

UMP UTP and CTP Nucleoside monophosphate kinase catalyzes transfer of Pi to UMP to form UDP;
Nucleoside diphosphate kinase catalyzes transfer of Pi from ATP to UDP to form UTP
CTP formed from UTP via CTP Synthetase driven by ATP hydrolysis
– Glutamine provides amide nitrogen for C4 in animals
UMP + ATP UDP + ADP
UDP + ATP UTP + ADP
nucleoside diphosphate kinase
CTP synthase (cytidylate synthetase)
N
N
O
H
O
Ribose 3 phosphate
N
NO
Ribose 3 phosphate
NH2glutamine +
ATPGlutamate +
ADP +Pi
CTPUTP
(in bacteria, ammonia donates the amino group)

Pyrimidine base Salvage
The major enzyme is pyrimidine nucleoside phosphorylase, which takes
the free base, + ribose-1-phosphate, to produce the nucleoside and free
phosphate. This enzyme has a very low affinity for thymine, so never
make a ribo-thyamine.
- Thymine phosphorylase will take thymine + deoxy-
ribose-1-phosphate to produce thymidine
This is different from purine nucleoside phosphorylase, which degraded
the nucleoside to the free base + ribose-1-phosphate
Once the nucleosides are formed, kinases will phosphorylate them
Uridine-Cytidine kinase thakes either uridine or cytidine,
plus ATP, to yield UMP or CMP plus ADP
Thymidine kinase takes thymidine plus ATP to make
dTMP plus ADP
Deoxycytidine kinase takes dC +ATP to produce dCMP
and ADP

This enzyme has low
affinity for thyamine,so
the thyamine
ribonucleotide is rarely, if
ever, produce
Pyrimidine base Salvage
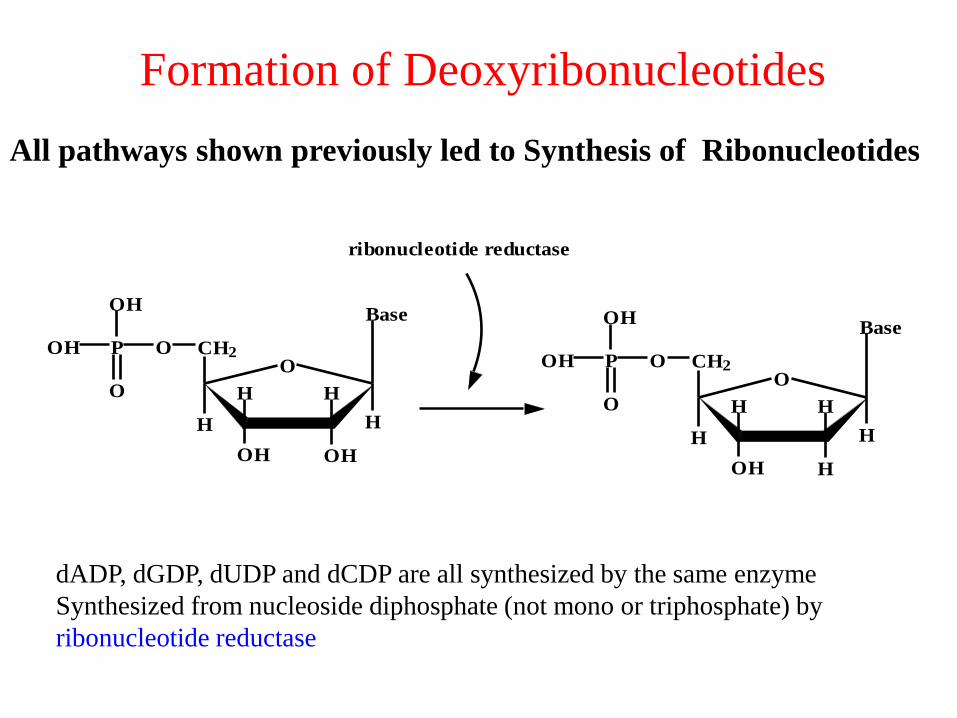
Formation of Deoxyribonucleotides
O
HH
OHOH
CH2
H H
OP
O
OH
OHBase
O
HH
HOH
CH2
H H
OP
O
OH
OHBase
ribonucleotide reductase
dADP, dGDP, dUDP and dCDP are all synthesized by the same enzyme
Synthesized from nucleoside diphosphate (not mono or triphosphate) by
ribonucleotide reductase
All pathways shown previously led to Synthesis of Ribonucleotides

Biosynthesis: Purine vs Pyrimidine
• Synthesized on PRPP
• Regulated by GTP/ATP
• Generates IMP
• Requires Energy
• Synthesized then added to
PRPP
• Regulated by UTP
• Generates UMP/CMP
• Requires Energy
Both are very complicated multi-step process which
your kindly professors do not expect you to know in detail

Purine Catabolism • All purine degradation leads to uric acid (but it might not stop
there)
• Ingested nucleic acids are degraded to nucleotides by pancreatic nucleases, and intestinal phosphodiesterases in the intestine
• Group-specific nucleotidases and non-specific phosphatases degrade nucleotides into nucleosides
– Direct absorption of nucleosides
– Further degradation
Nucleoside + H2O base + ribose (nucleosidase)
Nucleoside + Pi base + r-1-phosphate (n. phosphorylase)
NOTE: MOST INGESTED NUCLEIC ACIDS ARE DEGRADED AND EXCRETED.

Intracellular Purine Catabolism
• Nucleotides broken into nucleosides by action
of 5’-nucleotidase (hydrolysis reactions)
• Purine nucleoside phosphorylase (PNP)
– Inosine Hypoxanthine
– Xanthosine Xanthine
– Guanosine Guanine
– Ribose-1-phosphate splits off
• Can be isomerized to ribose-5-phosphate
• Adenosine is deaminated to Inosine (ADA)

Intracellular Purine Catabolism
• Xanthine is the point of convergence for the
metabolism of the purine bases
• Xanthine Uric acid
– Xanthine oxidase catalyzes two reactions
• Purine ribonucleotide degradation pathway is
same for purine deoxyribonucleotides

Adenosine Degradation

Xanthosine Degradation
• Ribose sugar gets recycled (Ribose-1-Phosphate R-5-P )
– can be incorporated into PRPP (efficiency)
• Hypoxanthine is converted to Xanthine by Xanthine Oxidase
• Guanine is converted to Xanthine by Guanine Deaminase
• Xanthine gets converted to Uric Acid by Xanthine Oxidase

AMP + H2O IMP + NH4+ (AMP Deaminase)
IMP + Aspartate + GTP AMP + Fumarate + GDP + Pi
(Adenylosuccinate Synthetase)
COMBINE THE TWO REACTIONS:
Aspartate + H2O + GTP Fumarate + GDP + Pi + NH4+
The overall result of combining reactions is deamination of Aspartate to Fumarate at the expense of a GTP
THE PURINE NUCLEOTIDE CYCLE

- The purine nucleotide cycle for
anaplerotic replenishment of
citric acid cycle intermediates in
skeletal muscle
- Occurs primarily in muscles
and brain (absent in most other
tissues).
- As AMP levels increase during
exercise, the cycle begins
-The ammonia produced can
buffer lactic acid, or can be
incorporated into glutamine
-Fumarate used as intermediate
for TCA

Uric Acid Excretion
• Humans – excreted into urine as insoluble
crystals
• Birds, terrestrial reptiles, some insects –
excrete insoluble crystals in paste form
– Excess amino N converted to uric acid
• (conserves water)
• Others – further modification :
Uric Acid Allantoin Allantoic Acid Urea Ammonia

Gout
• Impaired excretion or overproduction of uric acid
• Uric acid crystals precipitate into joints (Gouty Arthritis), kidneys, ureters (stones)
• Lead impairs uric acid excretion – lead poisoning
• Usually affect joints in the lower extremities (the big toe is the classic site)
• Xanthine oxidase inhibitors inhibit production of uric acid, and treat gout
• Allopurinol treatment – hypoxanthine analog that binds to Xanthine Oxidase to decrease uric acid production

Degradation of Pyrimidines
• CMP and UMP degraded to bases similarly to purines
– Dephosphorylation
– Deamination
– Glycosidic bond cleavage
• Uracil reduced in liver, forming b-alanine
– Converted to malonyl-CoA fatty acid synthesis for energy metabolism

Degradation of Pyrimidines
Note
1- Occurs in liver
2- End products soluble
(unlike purine degradation)
3- No associated disorders







![8. nucleic acid metabolism [compatibility mode]](https://static.fdocuments.net/doc/165x107/5875b2721a28ab8b618b6631/8-nucleic-acid-metabolism-compatibility-mode.jpg)










