
4 A. About Hibiscus cannabinus Linn. - Information and...
80
4 A. About Hibiscus cannabinus Linn. 1. Introduction Among the three species of Hibiscus, H. cannabinus Linn., H. sabdariffa Linn. and H. tiliaceus Linn., studied in the present investigation, H. cannabinus is economically the most important one. It is well known in the Western world as Kenaf. Although the term ‘Kenaf’ is Persian. The crop is probably originated in Africa (Hinsigi and Krisna, 1998). First time domesticated as early as 4000 BC in Western Sudan, it is widely cultivated as fibre crop in tropical and subtropical parts of the world. Besides India, major Kenaf growing countries are Argentina, China, Cuba, Egypt, Hawaii, Guatemala, Iran, Indonesia, Mozambique, North Africa, New Guinea, Peru, Spain, South Africa, Southern part of Zimbambwe, Thialand and Russia. Attempts have been made to cultivate this crop in Australia, Italy and USA for making pulp in paper industries (Charles-Edwards et al., 1983 and Webber and Bledsoe, 2002). Intercropping of the Kenaf is being practiced in India along with Jowar (Raji, 2008) and Ragi in Karnataka and Andhra Pradesh. There are different vernacular names for H. cannabinus L. in different languages. These are presented in Table 1. Table 1. Vernacular names of Hibiscus cannabinus L. Language Vernacular Names English Ambari hemp, Bastard jute, Bimli jute, Bimlipatam jute, Brown Indian hemp, Deccan hemp, Gambo hemp, Gombo hemp, Guinea hemp, Hemp hibiscus, Hibiscus hemp, Indian hemp, Java-jute, Kenaf, Mesta, Rosella hemp, Roselle, Siam jute, Thorny mallow, Wild stockrose, Hemp leaved Hibiscus. French Chanvre de Bombay, Chanvre du Deccan, Chanvre de Guinée, Chanvre de Gambo, Chanvre de roselle, Jute de Java, Jute de Siam, Kénaf, Ketmie à feuilles de chanvre (Belgium), Roselle. German Ambari, Dekkanhanf, Gambohanf, Hanfeibisch, Javajute, Kenaf, Rosellahanf, Roselle, Siamjute. Turkish Hibiskus
Transcript of 4 A. About Hibiscus cannabinus Linn. - Information and...

4
A. About Hibiscus cannabinus Linn.
1. Introduction
Among the three species of Hibiscus, H. cannabinus Linn., H. sabdariffa
Linn. and H. tiliaceus Linn., studied in the present investigation, H. cannabinus is
economically the most important one. It is well known in the Western world as Kenaf.
Although the term ‘Kenaf’ is Persian. The crop is probably originated in Africa
(Hinsigi and Krisna, 1998). First time domesticated as early as 4000 BC in Western
Sudan, it is widely cultivated as fibre crop in tropical and subtropical parts of the
world. Besides India, major Kenaf growing countries are Argentina, China, Cuba,
Egypt, Hawaii, Guatemala, Iran, Indonesia, Mozambique, North Africa, New Guinea,
Peru, Spain, South Africa, Southern part of Zimbambwe, Thialand and Russia.
Attempts have been made to cultivate this crop in Australia, Italy and USA for
making pulp in paper industries (Charles-Edwards et al., 1983 and Webber and
Bledsoe, 2002). Intercropping of the Kenaf is being practiced in India along with
Jowar (Raji, 2008) and Ragi in Karnataka and Andhra Pradesh.
There are different vernacular names for H. cannabinus L. in different
languages. These are presented in Table 1.
Table 1. Vernacular names of Hibiscus cannabinus L.
Language
Vernacular Names
English Ambari hemp, Bastard jute, Bimli jute, Bimlipatam jute, Brown Indian hemp, Deccan hemp, Gambo hemp, Gombo hemp, Guinea hemp, Hemp hibiscus, Hibiscus hemp, Indian hemp, Java-jute, Kenaf, Mesta, Rosella hemp, Roselle, Siam jute, Thorny mallow, Wild stockrose, Hemp leaved Hibiscus.
French Chanvre de Bombay, Chanvre du Deccan, Chanvre de Guinée, Chanvre de Gambo, Chanvre de roselle, Jute de Java, Jute de Siam, Kénaf, Ketmie à feuilles de chanvre (Belgium), Roselle.
German Ambari, Dekkanhanf, Gambohanf, Hanfeibisch, Javajute, Kenaf, Rosellahanf, Roselle, Siamjute.
Turkish Hibiskus

5
Polish Czyli kenaf, Ketmia konopiowata, Nacacha, nhacandora, canhamo brasilerio.
Sanskrit Manipuri, Sougri, Machika, Maryurika, Ambika, Sahasravatamulika, Sunishannaka, Nalita.
Hindi and Bengali Patsan, : Patsan,Bola, Chewla, Pitwa.
Marathi Ambadi, ambada.
Kannada Dirin da rani, Pundi.
Telgu Punti Koora, Goru, GonKura, Gaynaru.
Tamil Puliccha keerai, Gongura, Patumanji, Kasini.
Malyalam Attaparathi, Kanjaru.
Oriya Kanuriya
Gujarati Sheria, Ambari.
Source : Cooke (1967 a), Sastri, (1959) and Watt, (1890).
Hibiscus cannabinus has following synonyms
Abelmoschus verrucosus Walp., Hibiscus verrucosus Guill. and Per.
(http://www.plantnames.unimelb.edu.au/new/Hibiscus.html#cannabinus ).
2. Botany
It is a shrubby plant with green to reddish stem. Stem height ranges from 1.5
to 4 m (Siepe et al., 1997). Leaves are simple and compound, depending on the
variety and considered as varietal character (Singh, 2010). According to Joyner and
Pate (1956), there are two degrees of lobing in Kenaf. In some types lobing is deep
and a compound leaf structure is approached while, in other the lobing is very shallow
and an entire leaf from is maintained. Leaves are 2 inches across, glabrous, cordate
(those near the base often undivided) roundish ovate, the upper deeply palmately 3-5
lobed; lobes usually narrow–lanceolate, serrate, petioles1-2 inches long sometimes
prickly; stipules 3/8 inch long, subulate. Leaf also shows variation in attachment of
petioles with the stem. In some varities it is having angle of 900 while in other
varieties it is of 600-300. It is reported that in cannabinus varieties there is difference
in flower behavior. Flowers are large solitary, yellow in colour with reddish purple or
scarlet throat and appear in the month of January in the axil of leaf. Pedicels are very

6
short. Involucral bracts 7-10, free, 3/8 inch long, linear, acute, often with prickly
margins, shorter than the calyx. Calyx (in fruit) is 1 inch long, divided 3/4 of the way
down; lobes long, lanceolate, very acute, with strong midrib and thickened, often
prickly and with an oblong obovate gland at the base of each lobe. According to
Sivarajan and Pradeep (1996), H. cannabinus is characterized by a Calyx with whitish
arachnoid pubescence and also pustular, neither based on aculei, neither red nor
becoming fleshy. Corolla is yellow with purple at the center. Numerous stamens are
present, forming filamentous column surrounding the style, which is 17-23 mm in
length, dark red, with yellow or red anthers (Bukenya-Ziraba, 2004), Ovary is at
superior position, ovoid, villous, 5-celled and style shows 3-5 branches, which have
hairy arms with 2-4 mm length, each branch at the tip shows presence of capitate
stigma. Capsules are ovoid to cylindrical, beaked, and pubescent bearing from 18 to
20 seeds per capsule. The seeds are grey in colour (Singh, 2010). Seeds are reniform
to triangular large, grey to brown black, dotted with minute stellate scales (Cooke,
1967 a and Yadav and Sardesai, 2002).
Floral formula:
3. Anatomy
The anatomical studies of tap roots in 10 days old plant were performed by
Changdee et al. (2008). These studies were from basal part of the root and 5 cm from
the root tip. Tetrarch xylem was noticed, Casparian bands were quite prominent in
exodermis and endodermis. Although a large aerenchyma like intercellular space was
noticed in root cortex, it was not connected to the basal part of the root. Kenaf stem is
multicellular tissue which constitutes epidermal cell, libriform fibre cell, xylem fibre
cells, parenchymatous cell, wood ray cell, pit vessels, spiral vessels and trapezoidal
vessels. Among those fibre is the main cell. This cell delimited by presence of
primary wall (P) and secondary wall (S) in Kenaf bast and woody fibre cell types. The
secondary cell wall is divided into S1, S2 and S3 layers SEM and TEM analysis of
both core and bast fibres showed anatomical differences in three layers S1 layer is
cross spiral slightly thicker in bast and thinner in woody cells, S2 layer is
unidirectional spiral and widest among all fibres and slightly thicker wall with small

7
lumen, slender in appearance and few pits. However, woody fibres have thinner wall
with large lumen, stout in appearance and more pits (Chen and Shi, 1992). Individual
bast fibres are (15-) 2-3(-12)mm long and (7-) 15-25(-41)µm in diameter with a cell
thickness of 4-9µm (Bukenya-Ziraba, 2004). The anatomical studies of leaf surface by
Curtis and Lauchli (1987) revealed that epidermal cell thickness of abaxial surface
was 23.7 µm and adaxial surface was 28.0 µm respectively. These workers noticed
that the stomatal density of abaxial leaf surface was 218 mm-2 while, that of adaxial
leaf surface was 125 mm-2.
Datta et al. (1981) studied characteristics of mature embryo at 2 leaf
primordial stage. Root apex is of closed type. Procambia initiated in 2 loci, 1 near
cotyledon base, the other near the subterminal part of root, phloem started at 2 loci
(cotyledon and lower hypocotyl), xylem just below cotyledon base. Cotyledon
bundles had discontinuous differentiation. Primordial bundles differentiated
bidirectionally from base. There are different shapes of glandular trichomes on
adaxial epidermis of cotyledons and apical dome shaped shoot changed from almost
flat or concave to a conical in successive plastochroms after germination.
4. Cytogenetical Studies
For the identification of the plant at species level, chromosome number is
taken as basic characteristics. Diploid chromosome number of H. cannabinus is
2n=36 (where x=18) and it is considered as a lowest chromosome number in the
genus Hibiscus (Skovsted, 1941, Tjio, 1948 and Menzel and Wilson, 1961).
Autotetraploid H. cannabinus L. (2n =72) in which more than 50% of chromosomes
pair as trivalents or quadrivalents are also found. Cheng et al. (2002) analyzed 14
samples of Kenaf varities using RAPD markers in Japan. Their study revealed that
Kenaf can be divided into three major groups on the basis of characters, such as
middle stem diameter, whole stalk weight, and days to 50% flowering, but it is
difficult to identify individual varieties merely by the morpho-agronomic characters.
Thus it is clear that RAPD analysis is an effective tool in identifying Kenaf varieties
and determining their genetic relationships. Heterosis in Kenaf was studied by Pate
and Joyner (1958), Nelson and Wilson, (1965); Srivastava et al. (1978) and Patil and
Thombre, (1980). These studies revealed that three lines HC 625, HC 622, HC 602

8
were the best general combinations for the most of the important characters. Heterosis
for dry bark weight was highest in HC 602 versus HC 726. The highly significant F-
test values demonstrated the extreme of variability for general combining ability
(GCA) and specific combining ability (SCA). Chen et al. (2005) studied
developmental behaviour of gene expression in H. cannabinus L. with respect to plant
height and growth stages. They found that conditional and unconditional genetic
effects of plant height showed dominance while the additive effects were weak, also
plant height and stem diameter were not completely same. Genetic transformation in
Kenaf was attempted by Banks et al.(1993) and Srivatanakul et al.(2000). Transgenic
studies were also carried out by Kojima et al.(2004) with the help of well known
variety of Kenaf Aokawa No. 3’.
Singh (1988) attempted mutation breeding in Mesta for better quality. He
reported that induced mutants were more superior in fatty acids and amino acid
content. AFLP based identification and genetic relationships of Kenaf germplasm was
carried out by Cheng et al. (2004). Cheng et al. (2002) compared the genetic diversity
among Kenaf varieties based on the analysis of agronomic and RAPD data. Guo et al.
(2002) studied random amplified polymorphic DNA (RAPD) analysis among H.
cannabinus and related species. Samanthi et al. (2004) studied multiple shoot
regeneration from young shoots of Kenaf. These workers also performed light and
scanning electron microscopic analysis of benzyl adenine induced multiple shoot
regeneration in kenaf (H. cannabinus L.). Samanthi et al. (2005) studied factors
influencing Agrobacterium mediated genetic transformation of Kenaf.
5. Physiological Studies
a. Seed Germination
Kenaf seeds with 8 percent moisture retain viability for about five to six years
at 10 0C temperature (Singh, 2010). Seed storage becomes part of the Kenaf culture.
Kenaf seeds stored upto 4 years at 4 0C retained germination rates at ideal conditions
(Meints and Smith, 2003). Toole et al. (1960) reported that Kenaf seeds stored at 0 0C
remained viable with improved germination. Seeds stored at 8 or 12 % moisture could
show increase in germination rate almost 17 % after 6 months storage at 10 0C and
subsequently declined (Toole et al., 1960). However, Meints and Smith (2003)

9
reported that in seed lots stored at 10 0C did not show significant decline in
germination percentage. A soil temperature of 15 to 20 0C is most suitable for the
germination of seeds. Generally a sowing depth of 2.5 to 3 cm is suitable for good
germination. Soil moisture of 20-32 percent is good for the germination of seeds.
(Singh, loc. cit.).
The base temperature for Kenaf seed germination is reported to be 9.2 0C
(Angus et al., 1981), 9.7 0C (Carberry and Abrecht, 1990) and 8 0C (Angelini et
al.,1998). Carberry and Abrecht (1990) studied germination and elongation of
hypocotyl and radicle of Kenaf in response to temperature. They found that when
seeds of Kenaf c.v. Guatemala-4 were germinated in incubator at 8 constant
temperatures, final germination percentage was unaffected in the range of 15-35 0C
but it declined sharply at higher temperatures. The thermal time requirement of Kenaf
for 50 % seed germination has 8.2 0C dmm-1, the lag phase of hypocotyls elongation
required 17.0 0C d and linear hypocotyls elongation required 0.45 0C dmm-1.
Germination of oldest lot (1996) and youngest seed lot (2000) had the highest seed
germination percentage at 20 0C in January 2001 (Meints and Smith, 2003). White et
al. (1971) reported that seedling emergence was greater in chemically treated Kenaf
seeds than non treated seeds under field conditions, while, laboratory tests indicated
that such treatment had no effect on germination. The isolation, purification and
characterization of enzymes citrate synthase and lipase from seeds of kenaf has been
carried out by Zemlyankukhina et al. (1967) and Kausar and Akhtar (1979).
b. Growth
According to LeMahieu et al. (1991) Kenaf plant when grown in dense strands
showed unbranched habit and grow to a height of 8 to 14 ft. under favourable
conditions may reach upto 20 ft. Stem colour varies, it may be green to reddish or
puplish. Roots are with central single deep tap root along with wide spreading lateral
roots. Muchow (2009) studied relationship of population of Kenaf cultivar
Guatemala-4 and growth parameters under tropical conditions. The increased plant
density decreased all the growth parameters (stem length, basal stem diameter,
branching, number of leaves and nodes and leaf area per plant). At the end of growth
period Kenaf had attained a mean height of 2.47m (Ogbonnaya et al.,1998). Ching et
al. (1997) studied effect of gibberellic acid on the growth and yield of Kenaf. GA was

10
effective in increasing plant height and proved to be a potential plant growth
regulator. It is suggested by Arumingtyas et al. (2010) that a branching phenotype
may be produced due to the result of interaction between auxin and other hormones.
Branching is profuse in Kenaf plants before flowering, after which all the axillary
buds were converted to flower buds. A well grown and regularly irrigated Kenaf plant
showed 57 nodes. However, first flower bud was formed on 29th node, also long
internodes alternated with short internodes more vigourously after the 4th week of
growth. According to Wood et al.(1983), cultivars differ markedly in their response to
daylength but stem growth generally declines rapidly following the onset of
flowering. Leaf development varies in shape in different varieties from simple to
lobed. In Kenaf seedlings, first few leaves are not lobed. However, post juvenile
leaves are very deeply lobed (LeMahieu et al.,1991). Leaf characters are closely
related to growth and biomass productivity. This aspect was studied by Danalatos and
Archontoulis (2010) in cultivars Tainung-2 and Everglades 41 of Kenaf under
different agricultural inputs in Greece. They estimated the leaf area, specific leaf area
and Leaf area Index of above two cltivars of Kenaf. Three consecutive years
observation regarding leaf area Index showed that LAI values ranged from 4.2 to 6.4
at different sowing dates. However, with earlier sowing of the crop the higher LAI
values ranged from 3.9 - 6.8. The SLA during vegetative stages reached maximum
thereafter during flowering exponential decrease was noticed with decrease in day
length from13.5 to 10.94 (Danalatos, 1993). Biomasss accumulation showed steep
increase after 6th week of growth. Biomass in terms of leaf, root, stalk, shoot, wood
dry weight was increased with increasing age of Kenaf plant. According to Hossain
et al. (2011), the dry matter accumulation and partitioning in Kenaf varieties into
roots, stem and leaves varied substantially. Pioneer studies by Clark and Wolffs
(1969), revealed that growing season do interfere in chemical composition of Kenaf.
They further reported that hot water extractives and protein content were decreased
while; there was increase in lignin content and α-cellulose contents at 244 DAP than
90 DAP of Kenaf (Tainung-1 (T-1) cultivar). Rowell and Han (1994) studied changes
in chemical composition of Kenaf during normal growth. Ash, protein extractives, L-
arabinose, L-rhamnose, D-galactose, α D-mannose, contents decreased while lignin,
D-glucose and D-xylose content increased as the plant matured. Fibre length was

11
increased during early stages of growth than late stage of (mature) development. The
changes in phospholipids during growth of kenaf were studied by Tolibaev et al.
(1981).
c. Flowering and Photoperiodism
Flowering of most Kenaf cultivars is under photoperiodic control (Crane,
1947). Hibiscus cannabinus is a flowering plant sensitive to day length of 12.30 h
(Zen, 1982 and Singh, 2010). According to Carberry et al. (1992), Kenaf behaves as a
qualitative short-day plant remaining vegetative until daylength falls below 12.9 h.
Bukenya-Ziraba, (2004) indicated that flowering is influenced by the time of planting;
long days and high temperatures prolong the vegetative growth.The growing period
ranges from 110 to 140 days. Warner and Erwin (2003) reported effect of photo
period and daily light integral on flowering of five Hibiscus species (H. cannabinus).
Hinsigi and Krishna (1998) studied that H. cannabinus matures in 120 to 130 days.
According to ShivRaj (1978), 10 h light period is sufficient for flowering and mean
flowering time required in 69.22 days. Sowing dates determines the vegetative phase
of the crop in Kenaf. Early sowing expands the vegetative phase before the entry of
crop into critical photoperiodic levels. A drop in critical level of photoperiods
(arround12 h) that made crop to set into flowering. ShivRaj, (1978) reported that
plants treated with 10 h and 12 h photoperiod flowered in about 35 and 48 days
respectively as against 158 days required in control plants flowering is progressively
earlier if the short day treatments were interrupted less frequently or not at all. They
also suggested that the best time for sowing Kenaf for optimum vegetative growth is
late March to mid April. Vegetative period and the flowering time shortned with delay
in sowing. Kenaf sown in March and July flowered in mid to late September. Plants
sown in March and June were exposed to long days, high temperature, high humidity
and high rainfall which resulted in good vegetative growth. Kenaf flowers close
before noon on the same day. In view of Bukenya-Ziraba (2004), kenaf is mostly an
outbreeding plant, but upto 30 % self pollination occurs. According to Singh (2010)
Kenaf is a primarily self fertile, but it is often considered as a cross pollinated crop
because pollination sometimes takes place with the help of pollinating agents like
honey bees or insects.

12
d. Mineral Nutrition
It is admitted by ShivRaj (1978) that no detailed information is available about
mineral nutrition of Kenaf. Hansen (1981) studied nutrient content of Kenaf crop at
five growth stages. Samples of bark wood tops with foliage and seeds were analysed
for nitrogen, phosphorus, potassium, magnesium, calcium, zinc, iron, copper and
sodium. They noticed that the potassium content of above ground parts, throughout
the growth cycles was upto 280 kg ha-1. However, nitrogent content at two stages was
150 kg ha-1for initial and final stabilization at mature stage. Calcium content increased
from 65 kg ha-1to 105 kg ha-1with age. Zinc content was increased from 150 g ha-1to
350 g ha-1. At maturity Kenaf showed a higher content of iron i.e. upto 1400 g ha-1.
Hossain et al. (2011) studied nutrient partitioning in Kenaf varieties grown on sandy
bris soil. They noticed that the macro and micronutrient in Kenaf parts differed
significantly among the studied varieties. These workers also found that nitrogen
content is highest in leaves and lowest in stem, which was followed by K, Ca, P and
Mg. Kenaf gives good response to N, P and K fertilizers (ShivRaj, 1978). Hossain et
al. (2011) studied effects of N, P, and K at different levels on Kenaf growth and
photosynthesis. They noticed highest values of all the parameters studied such as
biomass, dry matter production, plant height, leaf number, root dry weight, stem dry
weight, photosynthesis and stomatal conductance at 200 N, 100 P and 100 K. The
values of these parameters decreased with increase in further concentration of N, P
and K. This might be due to imbalanced nutrient concentration or toxicity caused by
the higher nutrient concentration. Their study revealed that the profound effects on the
growth parameters such as leaf dry weight while, root/shoot ratio increased under N
deficiencies. In addition to this N, P and K deficiency also decreased plant height and
photosynthesis leading to lower biomass accumulation. Khader and Rama (1998)
studied mineral contents in two kenaf varieties Erragogu and Tellagogu at three
stages of plant maturity. They noticed that during plant maturity Ist stage (15 days) to
IInd stage (30 days) iron and manganese contents increased where as zinc and copper
contents were decreased as the plant matured. Investigation of the root tips of Kenaf
indicated that Cu and Zn mainly deposited in the cell wall, cytoplasm and nucleus. In
leaves the main sites of the deposition were chloroplast and nucleus, followed by cell
wall, cytoplasm and mitochondria. Vacuoles had the lowest Cu and Zn deposition.

13
High concentration of Cu and Zn depositions resulted in changes in root cell structure
such as cell wall deformation and vacuolization. However, cell wall deformities were
less in Kenaf (Yan, 2005). Anatomical studies of boron deficient plants by Hirai
(1950) revealed that, boron deficient plants showed disturbances in protoplasm and
cell wall and poor development of bast fibres causing wound gummosis. Banuelos et
al. (1996) studied response of Kenaf to boron amended water and soil. In the first
experiment they subjected kenaf plants to boric acid solution (containg 7.5 mg B l-1). In
the second experiment they raised kenaf plants in soil containg approx eq. 45 mg B kg-
1soil and extractable boron 7 mg B l-1 . They noticed 50 % and 27 % reduction in dry
matter over control in the first and second experiment respectively.These workers
estimated the leaf tissue B concentrations of kenaf ranged from 500 to 1400 mg B kg-
1DM during boron amended water irrigation and 422 mg B kg-1 DM when soil is
amended with boron. Banuelos et al. (1997) noticed appreciable accumulation of
Selenium (Se) in the leaves. Accumulation of Se can enrich the nutritive value of Kenaf
as leafy vegetable.
e. Gas exchange and Photosynthesis
Kenaf shows paracytic stomata having two subsidiary cells which lie at border
of the stomata, parallel with the long axes of the guard cells (Esau, 1977).
Archontoulis et al. (2005) estimated that at temperature above 35 0C a single leaf of
H. cannabinus may transpire the equivalent of 30 mm per day at full canopy. In three
kenaf varieties, G4, V36 and KK60 transpiration rates were compared. G4 had highest
and V36 had the lowest transpiration rates in all stages of growth except at 90 days
growth stage (Tahery, 2011). Some workers carried out field experiments and
observed stomatal opening during the night that can affect water requirement and leaf
transpiration (Muchow et al., 1980). This stomatal behaviour of kenaf explains the
night time evapotranspiration (Rosenberge 1969 and Cosentino et al., 2004). Reggi et
al. (2004) studied stomatal behavior and gas exchange in kenaf during night at two
different watering treatments (full irrigated and irrigated only at sowing). They
reported that both treatment leaves showed stomatal opening with increasing stomatal
conductance during late night and early in dawn period. An increased stomatal
transpiration during late night was noticed. Under all the artificial light treatments
there was unexpected increase in leaf transpiration under unirrigated treatment.

14
Kenaf is a typical C3 crop. Reddy and Das (2000) noticed that the primary
product of photosynthesis after 5 sec. of 14C assimilation was 3PGA. The
temperature optima of leaf photosynthesis was 32 0C and saturation irradiation
required for photosynthesis was 1600 µmoles m-2 sec-1. These workers further noticed
appreciable RUBISCO activity (245 µmole mg Chl-1 h-1) in Kenaf leaves and Km of
the enzyme activity for CO2 was 7-8 µM. Cosentino et al. (2004) indicated that Kenaf
has quite high net assimilation rate which contributes to high crop growth rates that
makes it superior in respect of biomass production and carbon sequestration.
According to them the C3 nature may lead to reduction in photosynthetic efficiency,
probably due to photorepspiratory losses at high thermal conditions. The net
photosynthesis during the whole measurement period in the best water conditions was
maintained at highest levels by the crop (28.4 µmoles CO2 m-2 s-1). Although there
was a negative correlation between net photosynthesis and leaf temperature, which is
irrespective of soil water conditions and the assimilation capacity reduced along with
increase in leaf temperature (Cosentino et al., 2004).
Tahery et al. (2011) noticed that net photosynthesis (A. net- assimilation
rate) in the three Kenaf varieties decreased with increasing age after 60 days stage.
However, at 60 days stage it reached a maximum values 6.5, 6.0 and 6.8 in G4, V36
and KK60 respectively. The variety KK60 maintained highest values for assimilation
rate than other two varieties and thus formed very efficient variety. Archontoulis et al.
(2005) reported maximum leaf assimilation rates (50 kg CO2 ha-1 d-1) in c.v.
Everglades-41 of Kenaf at global radiation exceeding 600Wm2. Kenaf maintained
appreciable photosynthetic capacity under salt stress (Curtis and Lauchli, 1986).
f. Stress Physiology
i. Water stress
Water Use Efficiency (WUE) of kenaf is studied by many workers (Banuelos
et al., 2002, Quaranta et al., 2000 and Danalatos and Archontoulis, 2010). It ranges
from 1.5 to 6.6 DM kg Water-1 for variety of soil regimes. Kenaf is being introduced
into arid regions (Francois et al., 1992) and increasingly been grown in other dry,
light textured and marginal soils with probability of water deficit developing during
growth. The experiment of Ogbonnaya, et al. (1998) revealed that leaf area, growth
and primary leaf production were equally affected and leaf number was reduced by 43 %

15
and/leaf area by 55 %, due to moderate water stress whereas, under severe stress
primary leaf initiation and leaf area development reduced by 66 % and 82 %
respectively. Branching was also reduced due to moderate stress by 75 % while, in
severe stressed plants branching was not observed. Flowering at 7th week growth
period was adversely affected by moderate stress and severe stress by 72 and 85 %
respectively. Flower buds which were formed before the onset of stress, withered and
dehisced. However, flower production recommenced after the plants were relieved of
stress. Number of nodes was also affected in moderate stress (41) and severe stress
(27) and reduced by 28 and 53 % respectively as compared to control (57 nodes).
Internode length was reduced with age in all treatments.
Biomass production was also significantly affected by water stress. There
were not many differences in the effects of moderate and severe stressed plants. At
the end phase, moderate stress reduced dry matter production of leaf, root, bark,
wood, stalk, shoot and total biomass by 74 % in moderate stress and 86 % by severe
stress. While, root-shoot ratio and bark-wood ratios were unaffected, however
stressed plants maintained a higher LAR throughout the experiment (Ogbonnaya, et
al., 1998). Transpiration rate in control plants decreased with maturity. However, in
stressed plants there was a steep fall and after rewatering transpiration rate rose
rapidly and in severely stressed plants transpiration went above the control and fall
soon afterwards. Leaf RWC was dropped with age in moderate and severely stressed
plants. At 2nd week stage, it was 76 % and 67 % in moderate and severe stressed
plants while, at last phase it was dropped to 58.57 % to 55.10 % respectively with
20.10 % and 24.80 % total reduction over control. Leaf water potential (χw) was
fairly affected due to water stress at beginning (Ogbonnaya, et al., loc. cit.).
The effect of water deficit on growth and water relation of Kenaf were
investigated by Ogbonnaya et al. (loc. cit.). They observed that water stress
significantly retarded vegetative growth in terms of plant height, collar diameter
growth, leaf development, branching, flowering and biomass accumulation. This
effect was also seen on biomass allocation in terms of root-shoot and bark-wood
ratios. According to these workers to overcome from drought, Kenaf roll its leaves or
bring about stomatal closure.
Ogbonnaya et al. (loc. cit.) investigated the effects of water deficit on the

16
physical and histochemical properties of Kenaf relevant to pulp and paper production.
Three watering regimes representing well watered control, moderate stress and severe
stress were imposed on the plants. Holistic analysis of the physical and histochemical
properties of Kenaf relevant to pulp and paper production indicated that water deficit
could improve the quality of pulp and paper produced from this plant.
According to Ogbonnaya, et al., (1998) Kenaf is found to possess high
desiccation tolerance capacity. This might be due to membrane resistance and /or
osmotic adjustment mechanism triggered when water deficit surpasses the critical
point of -0.5MPa. In contrast to this Kenaf is unable to employ earliness i.e. escape
mechanism in order to resist drought since flowering started at the same time in all
treatments. Another mechanism to protect plant from desiccation is closure of stomata
completely at the onset of drought though; this was at the expense of carbon
accumulation. A zero stomatal conductance in severely stressed plants on withhelding
watering exhibited sharp increase above the control upon rewatering (Ogbonnaya et
al. loc. cit.). By all these means, stomatal conductance and transpiration rates were
reduced completely. Kenaf is also found to roll its leaves during drought to avoid
water loss and drought.
Nkaa et al. (2010) studied growth performance of three Kenaf cultivars under
water stress in Eastern Nigeria. Kenaf cultivars Tainung 2, SF-459 and Everglades 41
were subjected to three water regimes well-watered control, moderate stress and
severe stress. The plants exhibited profound reduction in all vegetative aspects
including leaf development, branching, flowering and biomass accumulation. The
reduction in plant height was about 23 % and 39 % for moderately and severely
stressed plants respectively. Collar diameter reduced by 43 % in severe stressed stem,
Plant growth reduction was 27 % in moderately stressed plants. Water stress
effectively reduced plant growth thereby reducing the biomass production.
ii. Waterlogging stress
It is stated by Bukenya-Ziraba (2004) that Kenaf does not tolerate
waterlogging. A detailed study of responses of H. cannabinus to waterlogging stress
was carried out by Changdee et al. (2008, 2009 and 2010). These workers studied
influence of waterlogging on growth of Kenaf cultivar KK60 alongwith other two
cordage fibre crops H. sabdariffa and Corchorus olitorius. For this purpose these

17
workers designed two kinds of pot culture experiments. In one experiment (Changdee
et al., 2009) the treatment of waterlogging were as follows 45 days (105 DAS through
harvest), 60 days (90 DAS through harvest), 75 days (75 DAS through harvest) and
105 days (45 DAS through harvest) and well drained as control. In another
experiment (Changdee et al., 2010) a waterlogging treatment of 30 days was given on
30 DAS, 60 DAS, 90 DAS and 120 DAS while, no waterlogging treatment was
imposed on control plants. Their work revealed that Kenaf could grow better than H.
sabdariffa and jute under waterlogging conditions. It was noticed that 105 days
waterlogging caused 32.8 % reduction in plant height, 27.3 % reduction in stem
diameter 52.8 % reduction in leaf area, 51.2% reduction in leaf dry weight, 49.4 %
reduction in the shoot dry weight over the corresponding control plants of H.
cannabinus. Due to 105 days of waterlogging treatment the tap root length was
reduced by 52.4 % over the control. Hibiscus cannabinus subjected to waterlogging
treatment for 45, 60, 75, 90 and 105 days decreased fibre yield by 4.4, 7.3, 21.4, 28.6
and 40.8 of the control respectively. It was further noticed that a 30 days of
waterlogging treatments at 120 DAS caused 2 % reduction in fibre yield (Changdee,
2010). According to Changdee et al. (2008), fibre yield due to waterlogging in early
season was significantly decreased (13 % of the non-flooded control in Kenaf). The
anatomical studies of roots revealed that Kenaf formed the arenchyma in cortex of the
tap root under waterlogging condition. The formation of arenchyma in H. cannabinus
roots at seedling stage may contribute to the waterlogging tolerance (Changdee et al.,
2008). Hibiscus cannabinus seedlings formed exodermal Casparian bands throughout
from subapical to basal parts of the tap roots. Changdee et al. (2008) indicated that
Kenaf developed Casparian bands better than H. sabdariffa and jute in particular in
exodermis.
iii. Salinity
The screening of literature reveals that upto 1970 no attempt has been
probably made to study salt tolerance in H. cannabinus since there is no reference
cited in an exhaustive Bibliography ‘Plant Responses to Salinity: An Indexed
Bibliography’ published by US Salinity laboratory (Francois and Maas, 1978). Some
significant attempts have been made later to study the salt tolerance of this Hibiscus
species especially by Curtis and Lauchli. These can be summarized as follows.

18
Seed germination and seedling growth of Kenaf were studied by Liu and Li
(2011). Their study revealed that germination potential, germination rate, germination
index and vigour of Kenaf seeds were declined with increasing Na2SO4 and Na2CO3
salt concentration. Curtis and Lauchli (1985) noticed that seed germination of the
Kenaf cultivars were only slightly impaired by NaCl salinity upto 200 ml mol l-
1.These workers further noticed that dry weight accumulation after six weeks growth
was reduced 20-40 % by 75 mmol l-1NaCl (E.C. 7.8 dS m-1) and 70 -80 % by 150
mmol l-1 (E. C. 14.0 dS m-1). Vegetative development of three cultivars, C-108, G-45,
and E-71, and breeding line 15-2X, were similar under these salt treatments. Rate of
leaf emergence and leaf growth rate both declined linearly with increasing salt stress.
Francois et al. (1992) have studied salt tolerance of two H. cannabinus Linn. cultivars
viz. Everglades-41 and 7818-RS-10. Six salinity treatments of electrical conductivity
ranging from 1.1 to 6.0 dS m-1 were given. Vegetative growth, stem yield and fibre
length were measured. The mean dry weight yields of the stem during two years
experiments were reduced 11.6 % for each unit increase in soil salinity above 8.1 dS
m-1. Yield reduction resulted from both a reduction in plant height and stem diameter.
Increased salinity did not significantly affect fibre length. Based on these observations
these workers placed Kenaf in the category of salt tolerant plants. Kenaf behaves like
a typical moderate salt tolerant non-halophytic crop under salinity (Maas and
Hoffman, 1977). Biomass accumulation reduction at 75 mM salinity (9 dSm-1) was
45 % as with total leaf area (Curtis and Lauchli, 1986). Whole plant dry weight
accumulation was reduced to 45 % relative to control by 75 mM NaCl. However, in
37 mM NaCl, 14 % reduction was observed (Curtis and Lauchli, 1987). These results
were in agreement with those of earlier reports (Curtis and Lauchli, 1986) which
showed a threshold of about 37 mM NaCl for significant growth reduction due to
salinity. El-Katony, (1998) has studied effect of salinity-fertility interaction on growth
of H. cannabinus Linn. The plants were grown with three levels of macronutrients
(0.1 LA, 0.5LA, and 2 LA; LA= standard modified Long Ashton nutrient solution)
and three levels of salinity (0, 40 and 100 mM NaCl). The growth inhibition under
salt stress was most severe at adequate nutrient level (0.5 LA). Furthermore mild
salinity (40 mM NaCl) imposed at 0.1 LA was found to be beneficial. Curtis and
Lauchli, (1986) stated that under moderate salt stress there is slow cellular expansion

19
which limit leaf area in Kenaf due to reduction in cell number and epidermal cell size.
Curtis and Lauchli (1987) studied the effect of moderate salt stress on anatomy in H.
cannabinus Linn. (kenaf) and its relation to leaf area. They noticed that kenaf
responded to salt stress in a manner that was typical of moderately salt tolerant non-
halophytes. They observed that leaf area growth was more sensitive than leaf
emergence rate and dry matter accumulation. Leaf area and epidermal cell numbers
were prominently reduced while stomatal density increased with increasing salt stress
and leaf thickness was unaffected. Leaf area was reduced to 24 % at 37 mM salinity
and 40 % at 75 mM NaCl salinity; whole plant leaf area and leaf elongation rate
showed near by identical reduction in plants grown under similar conditions (Curtis
and Lauchli, 1985 and 1986). Epidermal cell size of cross sectional area of both
abaxial and adaxial surfaces wee reduced equally due to salinity (Curtis and Lauchli,
1987). There was significant increase in abaxial stomatal density with increasing
salnity. According to these workers the increase in stomatal density would tend to
counter the effect of partial stomatal closure in response to increasing tissue water
deficits and could help to explain the maintenance of high stomatal conductance,
measured on an area with increasing salinity in Kenaf (Curtis and Lauchli, 1986).
Findings of Curtis and Lauchli, (1987) support the hypothesis that anatomically leaf
area increase in Kenaf under salt stress shows the effects of internal water deficit
through a reduction in final cell size.
Curtis and Lauchli (1985) found that kenaf responded to salt stress excluding
Na+ from the shoot and partitioning Na+ and Cl- away from expanding leaf tissue.
Francois et al. (1992) noticed that excessive Cl accumulation occurred in leaf tissues
at high soil salinity levels. El-Katony, (1998) noticed that salinity caused reduction in
uptake, transport and accumulation of Ca in Kenaf. Salinity retarded uptake and
transport of Mg by H. cannabinus Linn. As a consequence of the differential effects
of salinity on Ca and Mg, the Ca/Mg ratio was altered. Banuelos et al. (1996) studied
accumulation in Selenium in Kenaf under increasing NaCl and CaCl2 salinity. They
noticed that generally there was a decrease in accumulation of Se in tissue with
increasing salt level.
Curtis et al. (1988) raised Kenaf plants for 35 days in solution culture at 1, 37,
and 75 mM NaCl under greenhouse conditions. Total carbohydrate increased in

20
mature and expanding leaves with increasing salinity. The majority of this increase
was as starch. Curtis et al. (1988) studied respiration in mature and expanding leaves
of kenaf exposed to moderate salt stress. They noticed that at 75 mM NaCl treatment
there was 29 % increase in respiration in mature leaves (1.75 (0.10) mM O2 m-2sec-1)
over control. According to them the net accumulation of non osmotically active
carbohydrates in expanding leaves suggests that growth was not limited by the
generation or availability of carbohydrates but rather by the ability of the plant to
effectively utilize this substrate in osmotic adjustment and growth. According to
Pervaiz and Sain (2003), net assimilation rate and CO2 exchange rate in kenaf were
insensitive to salinity. Salinity also affected turgor pressure through the decreasing
water potential showing a minimum osmotic adjustment in Kenaf (Curtis and Lauchli,
1985 and 1986).
6. Tissue culture studies
Cristofari et al. (1988) reported in vitro clonal propagation of H. cannabinus
L. In vitro studies of Kenaf were performed by McLean et al. (1992). They noticed
callus production along with caulogenesis and rhizogenesis from 4 weeks culture of
internodal stem explants of Kenaf. Internodal stem sections showed callus growth
and/or organogenesis with suitable auxin/cytokinin concentrations. MS-O
supplemented with NAA/BAP and 2, 4-D/kinetin could also serve as effective media
for callus production in Kenaf. However, both hormone combinations at various
concentrations were found beneficial for production of abundant callus in kenaf. It
was noticed that 83 % of the adventitious shoots produced roots when Kenaf explants
were placed on either media. It was further noticed that the kenaf shoot regenerated
from callus and adventitious organogenesis, from internodal stem explants.
Regeneration showed 3-13 % frequency depending on the plant growth regulator
combination. The survival frequency of kenaf transplants was 38 % without any prior
hardening. At the same time, it is admitted that organogenesis of kenaf via callus
culture was very poor and with very low regeneration efficiency and induction of
heritable mutants was possible (McLean et al., 1992). Reichert and Liu (1994 and
1996) described in vitro regeneration, protoplast isolation, culture and protoplast
fusion protocols for Kenaf.

21
A direct and simple regeneration using Kenaf shoot apex without a callus
phase was reported by Zapata et al. (1999). These workers noticed formation of single
shoot from a shoot meristem explants in their study. Concentrations of BA > 1 mg l−1
(4.4 µM) completely suppressed the shoot growth. Srivatanakul et al. (2000) also used
the shoot apex of the plant as an explant for generation of multiple shoots. One µmol
TDZ l-1 concentration in medium was found to be beneficial in inducing the highest
number of regenerated plants per explant from all the three cultivars (Tainung 1,
Tainung 2 and Everglades 71) studied. Two weeks duration was sufficient to induce
multiple shoots in axial shoot meristem of Kenaf. Further two week period with a
subculture (PGR free) is recommended for the induction of organogenesis i.e.
induction of roots in them. Histological studies of kenaf demonstrated de novo
regeneration of shoots from the shoot apex (Srivatanakul et al., 2000). Their study
confirmed shoot apex regeneration with two more kenaf cultivars 7N and SF-459
where there was induction of multiple shoots on the medium containing 1µmol TDZ l-1.
Chen et al. (2010) recently reported direct shoot organogenesis and plant
regeneration from cotyledonary node of Kenaf. Their study involved the use of
various plant growth regulators in the medium at various concentrations (5.0 mg l-1 of
BA, 0.3 mg l-1 IAA and 0.2 % (m v-1) F-68). After 21 days count of maximum number
of shoots per explant and frequency of shoot regeneration were 12.97 and 100 %
respectively. Among the three plant growth regulators, BA had greatest contribution
followed by IAA and F-68 in shoot induction. However, 0.5-2.0 mg l-1 IAA and NAA
were found effective in induction of root primordial in 14 days duration. The
maximum roots obtained with this combination were 13. In this respect IAA was
found more effective in induction of roots than NAA. According to Samanthi et al.
(2004), eight weeks were required to generate multiple shoots in young shoot explants
of all genotypes of Kenaf with significant differences among the different treatments
and cultivars. Addition of BA to the medium enhanced the shoot induction ability and
total number of shoots induced in Th3 and SF 459 was higher than those of BA free
medium. Kenaf (T2 and Th3) at 8.8 µM BA concentration showed highest number of
shoots (11explant-1). Kenaf regeneration efficiency was ranged from 42 to 99 %
among the different treatments, and increase in BA concentration resulted in reduced
shoot regeneration efficiency in all cultivars. Shoot elongation and root induction
22
were achieved simultaneously with no cultivar- dependent variation. Within a period
of three weeks about 96 % of the single shoot clumps were cultured on the medium.
No phenotypic variation was noticed between the seed grown and tissue cultured plants.
7. Phytochemical constituents
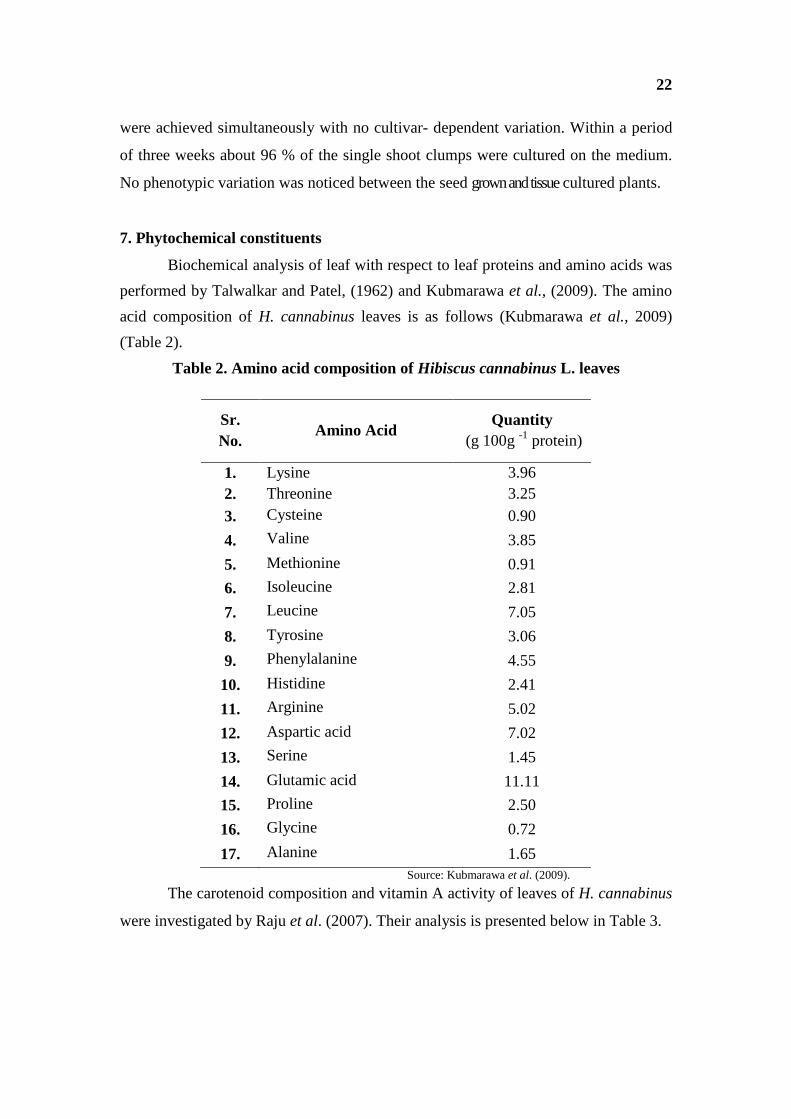
Biochemical analysis of leaf with respect to leaf proteins and amino acids was
performed by Talwalkar and Patel, (1962) and Kubmarawa et al., (2009). The amino
acid composition of H. cannabinus leaves is as follows (Kubmarawa et al., 2009)
(Table 2).
Table 2. Amino acid composition of Hibiscus cannabinus L. leaves
Sr. No.
Amino Acid Quantity
(g 100g -1 protein)
1. Lysine 3.96 2. Threonine 3.25 3. Cysteine 0.90
4. Valine 3.85
5. Methionine 0.91
6. Isoleucine 2.81
7. Leucine 7.05
8. Tyrosine 3.06
9. Phenylalanine 4.55
10. Histidine 2.41
11. Arginine 5.02
12. Aspartic acid 7.02
13. Serine 1.45
14. Glutamic acid 11.11
15. Proline 2.50
16. Glycine 0.72
17. Alanine 1.65 Source: Kubmarawa et al. (2009).
The carotenoid composition and vitamin A activity of leaves of H. cannabinus
were investigated by Raju et al. (2007). Their analysis is presented below in Table 3.

23
Table 3. Proximate analysis of carotenoids in the leaf of Hibiscus cannabinus L.
Sr. No.
Carotenoid Quantity (mg 100g-1 dry weight)
1. Xanthophylls
Neoxanthin 5.95
Violoxanthin ND
Leutin 33.97
Zeaxanthin 0.14
2. Total Xanthophyllsa 40.06
3. Provitamin A carotenoids
α-carotene ND
β-carotene 26.02
4. Total provitamin A carotenoidsb 26.02
5. Total carotenoidsc 66.08
6. REd 4338 Source: Raju et al.(2007). Detectable limit (0.1 p mol), RE=6 μg α-carotene and 12μ g β-carotene; a= sum of neoxanthin; violoxanthin, lutein and zeaxanthin; b= sum of α- and β- carotene; c= sum (mg 100g-1 dry weight) of total Xanthophylls and total provitamin A carotenoids; d=RE of α- and β- carotene and ND- non detectable.
Tkhin et al. (1981 a) investigated the dynamics of flavan compound
accumulation in roots H. cannabinus (Tkhin et al., 1981 b). These workers (Tkhin et
al., 1982) also reported occurrence of dimeric proanthocynidins. The proximate
analysis of Kenaf fibre indicated that it contains 18.9 % lignin 19 % pentosans and 66
% holocellulose (Hinsigi, 1989). Analysis of core from H. cannabinus bark showed
presence of various phenolic compounds such as acyclic phenylpropane lignanamides,
grossamide K and erythro-canabisine H, and the naphthol glucoside 2, 5-dimethyl-3-
O-beta-D-glucopyranosylnaphthol, along with six known compounds (Seca et al.,
2001 a). Four lignans, boehmenan H[2-(4-hydroxy-3-methoxyphenyl)-5-[3-(4-hydrox
y-3-methoxycinnamoyl oxy) propyl]-3-hydroxymethyl-7-methoxybenzodihydrof
uran], boehmenan K[2-(4-hydroxy-3-methoxyphenyl)-5-[3-(4-hydroxycinnamoy
loxy)-1-prope nyl]-3-(4-hydroxy-3-methoxycinn amoyloxymethyl)-7-methoxybenzo
dihydrofura n], threo-carolignan H [threo-1-(4-hydroxy-3-methoxyphenyl-2-[4-[3-
(4-hydroxy- 3-methoxycinnamoyloxy)propyl] -2- methoxyphenoxy] - 1, 3 –propan

24
odiol], and threo-carolignan K [ threo-1-(4-hydroxy-3-methoxyphenyl )-3-(4-hydroxy
-3-methoxycinnam oyloxy)-2- [ 4- [3- (4- hydroxycinnamoyloxy)-1-propenyl ]-2-
methoxyphenoxy]-1-propanol] as well as several other lignans aldehydes and a
tyramine derivative were isolated from the acetone extract of core of kenaf (Seca et
al., 2001a). Stipanovic et al. (2006) isolated and characterized (-)-3-Hydroxy-alpha-
calacorene from Kenaf through the technique of X-ray crystallography. Qualitative
studies of lignins in Kenaf were carried out by employing the method of pyrolysis-gas
chromatography technique in presence of tetramethyl ammonium hydroxide (TMAH)
to determine the ratios of abundance of Syringyl beta-aryl ether sub units to those of
the guaiacyl equivalents (S/G) in lignins. Syringyl beta-aryl/ guaiacyl ratio for in situ
lignins were obtained with an average of 3.1% relative standard deviation for H.
cannabinus (Kuroda et al., 2002). Lignin from the Kenaf are exclusively gamma
acylated with acetate group and forms beta-beta homocoupling and cross coupling
products of sinapyl alcohol is acylated at monomer stage and so it could be considered
as monolignol involved in the lignification reactions (Del-Rio et al., 2007). Two new
acyclic phenyl propane lignamide grossamide K and erythro-canabisine H, and the
naphthol, along with six known compounds were isolated from acetone extracts of
bark of H. cannabinus (Seca et al., 2001 b). Pascoal-Neto et al. (1996 and 1997)
studied the chemical composition of the macromolecules. The polysaccharides were
fractionated by successive extractions of holocellulose with aqueous KOH solutions.
The sugar composition was determined by hydrolysis of polysaccharides followed by
gas chromatography (GC) analysis of neutral sugars and spectrophotometric
determination of uronic acids. The results of general chemical analysis have shown
the different relative abundance of holocellulose, lignin, proteins, extractives and ash
in bark, core and foliage, at different stages of maturity. Pappas et al. (2002) isolated
and purified cellulose from Kenaf. These samples were characterized with the help of
diffuse reflectance infrared Fourier transform spectroscopy and 13C nuclear magnetic
resonance (13C-NMR) spectroscopy and the crystallinity was also determined. FTIR
spectroscopic analysis of pectins isolated from bark, wood and pith of four Kenaf
varieties showed 57 and 90 % degree of esterification and notably there was no
difference amongst the H. cannabinus varieties. Highest values of esterification was
noted in wood pectins (86-90 %) followed by pith (75-83 %) and lower in bark (57-64

25
%) (Pappas et al., 2004). The seed oil content in H. cannabinus is quite high. The
phospholipid composition of seeds of different cultivars of kenaf was investigated by
Tolibaev and coworkers (Tolibaev et al., 1976 and 1977 a b). The analysis of Kenaf
oil is carried out by Mohamed et al. (1995) and it is depicted in Table 4. It was
noticed in this analysis that Phosphatidyl choline, phosphatidyl ethanolamine, and
phosphatidyl glycerol were the dominant phospholipids.
Table 4. Seed oil content, lipid composition and fatty acid profile in Kenaf
Sr. No.
Component Quantity (%)
1. Oil content 21.4 to 26.4 2. Total phospholipids 3.9 to 10.3 3. Mean sterol percent 0.6 to 1.2 of the total oil 4. Sphingomyelin 4.42 of the total phospholipids 5. phosphatidyl ethanolamine 12.8 6. phosphatidyl choline 21.9 7. phosphatidyl serine 2.9 8. phosphatidyl inositol 2.7 9. lysophosphatidyl choline 5.3 10. phosphatidyl glycerol 8.9 11. phosphatidic acid 4.9 12. cardiolipin 3.6 13. β-sitosterol 72.3 of the total sterols 14. campesterol 9.9 15. stigmasterol 6.07 16. Palmitic acid 20.1 of the total fatty acids
17. Oleic acid 29.2
18. Linoleic acid 45.9
19. Palmitoleic acid 1.6
20. Linolenic acid 0.7
21. Stearic acid 3.5
22. Medium (C12---C14) and long (C22---C24) chain fatty acids
˂1
23. Eight unidentified phospholipids - Source: Mohamed et al.(1995) and Bukenya-Ziraba, (2004).
The composition of essential oils was studied (Kobaisy et al., 2001) with the
help of GCMS examination technique and Fifty-eight components were characterized
with (E)-phytol (28.16 %), (Z)-phytol (8.02 %), n-nonanal (5.70 %), benzene

26
acetaldehyde (4.39 %), (E)-2-hexenal (3.10 %), and 5-methylfurfural (3.00 %) as the
major constituents.
8. Cultivation
Hibiscus cannabinus can successfully grow at the height of 3000 ft above sea
level and at latitude of 450 N to 480 N in Russia and that to in South Africa at latitude
of 300 S. Although, the crop is affected by frost and extreme waterlogging, it shows
wide adaptability to soil and climatic conditions. It prefers warm humid climate, rich
loamy soil and about 50-90 cm rainfall. The crop is generally sown in India in late
July or early August in Kharif season. The seeds are either broadcasted or drilled. A
spacing of 25 to 30 cm between rows and 7 to 10 cm between plants is optimal for
good yields. In case of broadcasting sowing, plant to plant spacing is maintained at
12-15 cm by thinning. Generally a sowing depth of 2.5 to 3 cm is favourable for good
germination. A seed rate of 15 to 17 kg ha-1under broadcast and 13-15 kg ha-1 in line
sowing has been recommended. Irrigation is given to the crop during sowing time.
The crop gives good response to fertilizer application. Application of inorganic mixed
fertilizers (N, P and K) at the rate of 40:20:20 in split dose is optimal for higher fibre
yield. Organic fertilizers in the form of compost are also applied to field at the rate of
4-5 tones per ha. Two weeding are generally better, first at three week stage and
second at five week stage. Chemical weed control is also effective (Webber, 1992).
Generally harvesting is done with the help of sickle by cutting the 4 m tall plants close
to the ground. Kenaf has many varieties cultivated and suitable for different countries.
Most of the countries of the World have bred their own varieties suitable for their
local needs. Some of the important varieties of H. cannabinus (Kenaf) in India are as
follows (Singh, 1988):
· HC-583: This is a selection from a material collected from Nigeria. It was
developed at CRIJAF, Barrackpore. The plants are green with irregular light
flush of red pigment. The leaves are entire and cordate. The plants flower in
150-180 days after sowing. The flowers have yellow petal with red stigma. The
seed colour is greyish black. The seeds are sown in the month of April and it is
suitable for growing in the states of West Bengal, Assam and Orissa. The
variety can give fibre yield of 25-30 q ha-1.

27
· AMC-108: This variety is the outcome of selection of indigenous material and
was developed by the Agricultural Research Station, Amadalavalasa (A. P.).
The plants have light red pigmentation which extends even to the petiole. The
leaves are deeply lobed (5-7 lobes) and slightly red pigmented. It takes 150 days
to mature for harvest. The seeds are greyish black in colour. The variety is
suitable for sowing during April-May in the northern region of the country,
(Bihar and Orissa). The fibre yield is upto 25-30 q ha-1.
· HC-269: The variety has been released from CRIJAF Barrackpore. It is a
product of selection process from a local material. The plants are green with
irregular flush of red pigment. The leaves are entire and green. The variety
matures within 145-160 days for fibre purpose. The seeds are grey and
subremiform in shape. The seeds can be sown in April and can yield upto 20-25
q ha-1 of fibre.
Many diseases are reported on H. cannabinus (Singh, 2010).
ü Anthracnose – It is caused by fungus Colletotrichum hibisci (syn.-
Volutella). Affected part shows patches, and wilting of plant takes place.
Defoliation takes place, Flowers and seed capsules are damaged. It is
controlled by spraying copper oxychoride at the rate of 3 kg ha-1.
ü Tip rot – It is caused by Phoma spp. (syn. - Trichosphaeria spp.). Tip
browning of the growing plants, stipules, and young leaves, leaf buds etc.
is observed. For the better recovery, copper fungicide is effective.
ü Root rot/collar rot –It is caused by Rhizoctonia bataticola alone or in
combination with Fusarium oxysporum. The plants show wilting and
finally die. Mercurial compounds or captan are recommended to control it.
ü Eye rot – It is caused by Myrothecium roridum. The infected plant shows
eye shaped patches on the stem and leaves. Necrotic spots on stem are
initially smaller later on spots elongate and become brown and infected
stem is broken. Copper fungicide helps in controlling the disease.
Tobacco Necrosis Virus (TNV) and Hibiscus Latent Ring Spot Virus
(HLRSV) also attack the plant (Bukenya-Ziraba, 2004). Leaf curl disease of Kenaf
(H. cannabinus L.) in India has been found to be associated with begomovirus and
beta satellites (Chatterjee and Ghosh, 2007). Seventeen different species of insects

28
were detected on kenaf plant, out of which cotton Flea beetle Podagrica punctiollis
Weise is of negative economic importance and most serious at early seedling stage
(Bukenya-Ziraba, 2004). According to Bukenya-Ziraba (2004), kenaf is susceptible to
root-knot nematodes that reduce growth and yield on lighttextured soils. Root Knot
disease in plant is mainly caused due to infestation of either Meloidogyne javanica or
M. arenaria. Paul et al. (2006) reported that, in nematicide treated and untreated
fields with soil naturally infested with either Meloidogyne javanica or M. arenaria,
root rot indices indicated that Kenaf c.v. Everglade 71, baseline j-1-113 are resistant
verieties.
9. Uses
a. Human Nutrition
As a vegetable H. cannabinus is widely grown in Africa (Bukenya-Ziraba,
2004). The tender leaves are consumed as vegetable in many parts of India (Tanaka,
1976). Leaves are a delicacy and used in sausages in southern part of India. Leaves
and petiole contain 15 to 30% crude proteins with high digestibility. The protein yield
of a Kenaf crop in estimated to 400 to 500 kg ha-1 (Webber and Bledsoe, 1993). In
some parts of Andhra Pradesh a Pickle of Kenaf leaves under the name Gongura is
prepared. The composition of Kenaf leaves is as follows
Table 5. Chemical composition of Hibiscus cannabinus leaf.
Sr. No. Component Quantity
1. Water content (%) 79.0 2. Protein content (%) 5.5 3. Crude protein content(%) 13.78 4. Lipids(%) 2.33 5. Fat (%) 1.2 6. Carbohydrate (%) 12.2-37.67 7. Crude fibre %) 2.3-29.61 8. Energy 280 kJ(67 KCal) 9. Ash content (%) 5.11 10. Ascorbic acid (mg100g-1) 75 11. Ca (mg100g-1) 484 12. P (mg100g-1) 18 13. Fe (mg100g-1) 12.1 14. Phytic acid (mg100g-1) 19.78 15. Tannins (mg100g-1) 2.74

29
16. Oxalate (mg100g-1) 158.5 Source: Leung et al.(1968), Bukenya-Ziraba (2004) and Kubmarawa et al.(2009).
Ibnusaud and coworkers were granted US Patent (US 6127553 A) in the year
2000 for developing a process for isolation of Hibiscus acid ((+) hydroxyl citric acid
lactone) from leaves of H. cannabinus L. Earlier in 1972 Shchiparev and Soldatenkov
investigated the metabolism of this organic acid in leaves of kenaf (H. cannabinus-
vulgaris).
b. Forage Kenaf is important forage crop, which can tolerate multiple cuttings. Multiple
harvests produced linear stems more palatable foliage and yield 0 to 65 t ha-1 forage in
three harvests. The stem tip material has potential as animal feed. The plant tops are
with high digestibility and used as cattle and sheep feed (Killinger, 1969, Swingle et
al.1978 and Hays, 1989). According to Muir (2002), Kenaf has shown promise as a
forage when it was grown in semiarid regions.
Chantiratikal (2005) reported fodder value of Kenaf with respect to nutritive
value, fodder proteins and concentration of fibre in diet of cattles. He has also
suggested that kenaf can serve as a substitute for alphalpha hay and soybean meal
based on its comparative dietary composition
Trang (2004) investigated some aspects in two Kenaf verities (HC-K465/118-
K465) such as rate of growth of root system, crude protein content, dry matter yield
and evaluated cattle feed value and palatability score. He found that Kenaf plants
could grow well in constructed wetlands and could serve as a source of animal feed.
c. Fibre
Hibiscus cannabinus is largely cultivated for its fibre which is extensively
employed by the natives in the manufacture of rope, coarse sacking and other articles
required for agricultural purposes (Cooke, 1967 a). It has long been used in trade of
cordage products in making twines, ropes, sacs, and fishing nets. World production of
Kenaf fibres is estimated at 400000 tons per year, India being the largest producer
(Bukenya-Ziraba, 2004). Kenaf is used as a part of interior material of car such as
head liners and automobile dash boards. It would be a natural substitute for fibre glass
in future. It serves as material for carpet padding and corrugated medium and fire

30
resistant differential density Kenaf particle boards (K Boards). It is also used in
synthetic fibres molded plastics (Hinsigi and Krishna, 1998). PLA-based (Polylactic
acid) materials are a new class of materials that in recent years have aroused an ever
growing interest due to the continuously increasing environmental awareness
throughout the world. They essentially consist of synthetic polymers based
composites reinforced with natural fibres or other micro or nano fibre. According to
some workers, Kenaf is one of the mostly studied sources of natural fibre
reinforcement for PLA (Huda et al., 2008 and Garcia et al., 2008).
Kenaf is a bast fibre obtained from the stem of the plant. Stem height varies
from 1.5-4 mss. The stem presents a central core rich in short fibres and an external
bark with long fibres. On a dry weight basis, the bast fibre content of the stem ranges
from 21% in wild accessions to 36 % in modern cultivars (Bukenya-Ziraba, 2004).
During growth cycle of the plant, the bast fibre length increases first then decreases. It
again increased at final stage of growth (Han et al., 1995). This trend may be related
to several factors as indicated by Clark and Wolff (1969) alongwith cell wall (Rowell
and Han, 1994). The fibres are generally obtained by the process of retting. The
properties of fibre are partly dependent on the process of retting (Danladi 2008). In
view of Chee Hong (2004), biologically retted fibre composites showed better
physical and mechanical properties compared to chemically retted fibre composites at
the same fibre content and lay-up method. These fibres are strong and tight fibres with
continuous length that could be cut to the desired length for either spinning or
composites formation. There are two types of fibres, long bast fibres and short wood
fibres. The proximate analysis of Kenaf fibres is presented in Table 6.
Table 6. Proximate analysis and fibre dimensions in Kenaf
Sr. No.
Fibre dimension Quantity
1. Cold water solubility (%) 4.56
2. Hot water solubility (%) 6.42
3. Alcohol benzene solubility(1:2v v-1) (%) 4.28
4. 1% NaOH solubility(%) 28.50

31
5. Lignin (%) 18.9
6. Pentosans (%) 19.0
7. Holocellulose (%) 66-71.80
8. Hemicellulose (%) 25.05
9. α-cellulose (%) 46.75
10. β-cellulose (%) 10.24
11. γ-cellulose (%) 11.50
12. Ash (%) 1.56
13. Silica (%) 0.35
14. Av. Fibre length (mm) (l) 1.0
15. Fibre diameter (mm) (d) 0.024
16. Aspect ratio (l/d) 41.6
17. Burst factor 28.8
18. Tear factor 85.2
19. Breaking length (m) 50.17 Source: Hinsigi, (1989) and Dutt et al.(2009). Holocellulose: Hemicellulose+cellulose; d= diameter; l= length
d. Paper and Pulp Industry
Kenaf is attracting a considerable interest of the paper industry as a source of
good quality cellulose fibres (Siepe et al., 1997). Being an annual herbaceous plant, it
is fast growing and high yielding and pulp is easily obtained (Clark et al., 1962).
According to Angelini et al. (1998), kenaf can be used as an alternative raw material
for pulp due to its excellent advantages of being renewable, inexpensive and growing
ability under severe conditions such as low water supply and little fertilizer.
According to Francois et al. (1992) kenaf is 3-5 times more productive per
unit area than pulpwood trees producing pulp with quality equal to or superior to that
of many woody species. In Italy several studies have identified Kenaf to be an
interesting substitute for woody species utilized for paper pulp production (Petrini et
al., 1994). Ashori et al. (2008) have tried to improve the surface property of

32
handsheets prepared from the Kenaf material, and noticed that a bio-polymer of
chitosan has ability to form films that improve the surface properties of paper when it
is applied to the surface of the sheet and for preparation of net to trap the animals.
e. Medicinal Uses
i. Role in Human Health
From ancient times, H. cannabinus Linn. has been used as a folk medicine in
India and Africa for the treatment of blood and throat disorders, to check excessive
secretion of bile leading to biliness or longestion of liver with acidity, bilious
conditions, fever and puerperium, stomach problems, earache and used as an
appetizer. It is also used to cure dysentery, pains and bruises (Lee et al. 2007).
El-Basheir and Fouad, (2002) studied the effect of a mixed cream prepared
from Lawsonia alba L. (Henna). Trigonella faemum-gracanum (Fenugreek), H.
cannabinus (Hibiscus) and Artemisia cina (Wormseed) plants, on one hundred lice
infested patients (90 females and 10 males) with different ages and hair length. They
noticed that there was complete disappearance of head lice within a week due to
cream application.
ii. Pharmacological Studies
Antimicrobial and cytotoxic activities of six lignans isolated from the core
of bark of acetone extract of core of bark of H. cannabinus have been investigated.
Two compounds (not characterized) are strongly cytotoxic against Hela, Hep-2 and
A-549 Cell lines shared moderate activity (Moujir et al., 2007). Cell viability testing
experiments were carried out using a human leukaemia cell-line HL60 for in vitro
phytotoxic activity of 93 terrestrial species with about 155 extracts, in Penninsula
Malaysia (Ong et al., 2009). These workers noticed that among these species 29
species including H. cannabinus were able to reduce the in vitro cell viability by more
than 50 % when exposed to 9.6 J/cm(2) of a broad spectrum light when tested at a
concentration of 20 µg ml-1. Shivali et al. (2010) studied antihyperlipidemic effect of
hydroalcoholic extract of Kenaf leaves in high fat diet fed rats. They noticed that the
extract exhibited a strong dose dependent anti hyperlipidemic activity and effective in
reducing the levels of serum TC, TG, LDl-C and TBARS. The extracts effectively
prevented the liver microvesicular steatosis in hyperlipidemic rats. Essential oil
fractions of kenaf showed phytotoxic and fungitoxic activities (Moujir et al., 2007 and

33
Lee et al., 2007), H. cannabinus aqueous extract was found to be haemantically
active and anti-oxidative. It also exhibits hepatoprotective activity against carbon
tetrachloride and paracetamol induced liver damages in rats. Leaf extract in 80 %
ethanol caused immunomodulatory effects in activated macrophages (Lee et al.,
2007).
f. Biomass
According to Cosentino and Copani (2003) and Alexopoulou et al.(2005),
Kenaf due to its property to yield high biomass and elevated fibre content can be used
as a biomass for energy for its potential role in agroecosystems involving biomass
production as substitute for non-renewable resources. Baldwin et al.(1996) estimated
that Kenaf biomass yields ranged from 14 to 22 mg ha-1. Kenaf biomass was taken by
Meints and Smith (2003) on field plot by harvesting four rows equivalent to 0.006 ha.
The biomass yield was expressed on whole plant mass on dry weight basis. Their
experiments revealed that the biomass yields of Everglades 41 (E41) variety of kenaf
ranged from 12.39 to 14.57 mg ha-1 in 1999 and 16.82 and 18.47 mg ha-1 in 2000, but
were not different between storage duration. Late maturing cultivars of Kenaf, yield
potential stem biomass which usually has high non-grain biomass production at
central Italian latitudes (Benati et al., 1990 and Siepe et al., 1997).
g. Phytoremediation
Phytoremediation is one of the promising strategies to remove metals from
contaminated soil with plants and it is a simple, low-cost, and environmentally
friendly procedure. Among the species tested Hibiscus cannibinus was found capable
of Boron phytoextraction in soils containing from 1 to 10 mg kg–1 of B (water
extracts) and were able to reduce up to 24 % of B content in the soil in sixty months
(Banuelos et al.,1996).
The studies of Pais and Jones (2000) and Kabata-Pendias and Pendiais
(2001) also indicated that H. cannabinus is a potential candidate for phytoremediation
of lead (Pb). Ho et al. (2008) found that lead was totally absent in the leaves but 85 %
of the total plant ‘Pb’accumulated in roots. Munusamy and Agamuthu (2012) recently
observed sequestration of 0.06 to 0.58 mg Arsenic and 66.92 to 461.72 mg iron per g
plant weight in Kenaf roots and indicated that due to such high ability to tolerate these
metals and avoid phytotoxicity. Takahashi et al. (2008) showed that an ambient

34
concentration of NO2 in Kenaf avoids Cd. Hibiscus cannabinus when labeled with
NO2 fumigated for 8h and uptake and assimilation was determined by mass
spectrometry and Kjeldahl-nitrogen based mass spectrometry (Takahashi et al., 2005),
it was found that the plant performed a high uptake and assimilation during day time
as compared to night (Day 1100 to 2700 ng N mg-1 DW). Kenaf showed night uptake
and assimilation as high as 1500 ng N mg-1 DW comparable with CAM plant Aloe.
This experiment indicated that Kenaf is a potent phytoremediator of NO2 both during
day time and at night.
h. Oil
Seeds of H. cannabinus are economically important and serve as a source of
oil. Kenaf seed has 17.22 % fatty oil. This oil is used by soap making industries in all
parts of the world. The oil can be used in preparation of lubricants, linoleum, paints
and varnishes. Kenaf's relatively high oil content and its similarity to cotton seed oil
suggest that the seed oil may be used as a source of edible oil (Mohamed et al., 1995).
It is edible oil but it has minor antinutritional factors such as cyclopropene and epoxy
fatty acids. For cooking purpose, it has to be subjected to hydrogenation and it is to be
refined. It is roughly estimated that 50000 t of Kenaf oil is produced every year,
valued at Rs-100 Crores (Hinsigi and Krisna, 1998). This oil has following
characteristics.
Table 7a. Characteristics of Hibiscus cannabinus seed oil
Sr. No.
Oil Characteristic Quantity
1. Fat content (%) 18-20 2. Specific gravity 0.917-0.926 (150/150) 3. Acid value (%) 0.5-10 4. Saponification value 187-190 5. Iodine value 90-105 6. Refractive index at 400C = 1.465 q
Source: Sastri, (1959) and Singh, (2010).
The oil cake remaining after oil extraction contains following ingredients
(Bukenya-Ziraba, 2004 and Singh, 2010).

35
Table 7b. Proximate analysis of Hibiscus cannabinus oil cake.
Sr. No.
Component Quantity (%)
1. Moisture 9.26
2. Crude protein 32
3. Crude fibre 8
4. Oil 9.76 Source: Bukenya-Ziraba, (2004) and Singh, (2010).
The above composition suggests that the oil cake may serve as animal feed.
i. Other Uses
Kenaf can be used as a substitute for fibre glass, filtration media making and
food and bedding material for animals (Kugler, 1996 and Sellers and Reichaert,
1999). Bark, wood and pith extracts were utilized for isolation of pectins. Cuba 108
variety of H. cannabinus was utilized for isolation of pectins (Pappas et al., 2003).
Juhaida et al. (2010) manufactured polyurethane (PU) liquefied kenaf core (LKC)
with molecular weight (MW) of 2666, viscosity of 5370 mPa and solids content of
86.9 % and it is widely used as a wood laminating adhesive. It was noticed by
Kobaisy et al., (2001) that the seed oil had antifungal activity against Colletotrichum
fragariae, Colletotrichum gloeosporioides, and Colletotrichum accutatum but it
exhibited little or no algicidal activity.
B. About Hibiscus sabdariffa Linn. 1. Introduction Hibiscus sabdariffa Linn. var. sabdariffa known as Lal Ambari in India but
more popular under the name roselle is one of the noteworthy species belonging to
genus Hibiscus. The species was first described by Flemish botanist M. de L’Obel.
According to McClintock and El-Tahir (2004), the species probably originated in
Africa and was perhaps first domesticated in Sudan about 6000 years ago. At the
same time, some workers consider it to be a native of Asia (India to Malaysia). At
present, the plant is cultivated in different Asian and African countries. In India
roselle is cultivated in various parts of Punjab, Uttar Pradesh, Andhra Pradesh,

36
Assam, Bihar, Madhya Pradesh, Maharashtra and West Bengal (Gautam, 2004). The
wide cultivation range indicates wide adaptability of this species to variety of climatic
conditions. On the basis of growth habit and mode of utilization H. sabdariffa is
broadly classified under two varieties H. sabdariffa L. var. sabdariffa and H.
sabdariffa L. var. altissima Wester. Hibiscus sabdariffa is a multipurpose species
yielding vegetable, fibre and economically important red calyx. The species has also
appreciable medicinal value and numbers of pharmacological studies have been
carried out.
There are different vernacular names for H. sabdariffa L. in different
languages. These are presented in Table 8.
Table 8. Vernacular names of Hibiscus sabdariffa L.
Language
Vernacular Names
English Indian sorrel, Jamaica sorrel, Red sorrel, Roselle. Java jute, Karkade, Red tea, Royal roselle, Florida cranberry, Rozelle, Guinea sorrel, Sour-sour, Queensland jelly plant (Aust.), Jelly okra, Fibre Hibiscus, Chinese lantern, Coral Hibiscus, East-African Hibiscus, Fringed Hibiscus, Fringed rose-mallow, Japanese Hibiscus, Japanese lantern, Skeleton Hibiscus, Lemon bush, Jamaican sorrel, Rosella, Guinea sorrel, Sour-sour, Jamaica sorrel.
Sanskrit Manipuri, Sougri, Machika, Maryurika, Ambika, Sahasravatamulika, Sunishannaka
Hindi Lal-mista, patwa, chukar,Lal-ambari.Patwa, Gongura
Assam Chukiar
Bengali Lal-ambadi, patwa, Lal-mista, Chukar.
Marathi Tambadi-Ambadi, Lal ambadi, Patwa. Kannada Palachakiri, Pundibija Telugu Yerra gogu
Tamil Pulichchai Kerai, Gogu
Malyalam Polenchi, Pulichchai
Oriya Kanuriya
Gujarati Sheria
Polish Hibiskus szczawiowy, Ketmia szczawiowa.
French Chanvre de Bombay, Chanvre du Deccan, Chanvre de Guinée, Chanvre de Gambo, Chanvre de roselle, Jute de

37
Java, Jute de Siam, (Belgium), Roselle, Oseille de Guinée, Thé rose d'Abyssinie, Oseille de Guinée.
German Malventee, Rosella, Afrikanischer Eibisch, Hibiscus-Tee, Karkade-Tee, Roselle, Rote Malve.
Dutch Surinam, zuring
Portuguese Vinagreira, Azeda de Guine, cururu azedo and quiabeiro azedo.
African Karkade or carcade Source:Gautam, 2004 and Mahadevan et al.2009.
Hibiscus sabdariffa has some synonyms, these include Abelmoschus cruentus
Bertol, Hibiscus digitatus Cav., Hibiscus gossypiifolius Mill., Hibiscus sanguineus
Griff., Sabdariffa rubra Kostel and Hibiscus rosa-sinensis L. var. schizopetalus Mast
(http: //www. plant names. unimelb. edu. au/new/ Hibiscus. html# cannabinus,
Multilingual Multiscript Plant Name Database Sorting Hibiscus names)
2. Botany
The plant is erect, glabrous and unarmed and attains a height of about 3 to 3.5
m. having basal diameter of 1.0 to 2.0 cm (Singh, 2010). It has deep penetrating tap
root (Duke, 1983). Stem is with branches and is purple. Hibiscus sabdariffa species is
broadly classified under two varieties H. sabdariffa var. sabdariffa and H. sabdariffa
var. altisimma Webster (Mahadevan et al.,2009), former is generally bushy and
pigmented and cultivated for its edible calyces and the latter comprises tall growing
unbranched types cultivated for the stem fibre, roselle. It is classified into four main
groups on the basis of presence of pigmentation on the stem. They are full green,
green pigmented, green light red and red (Singh, 2010). Leaves are compound 2-3
inches long, usually 3-5 deeply lobed, cuneate at the base. Sometimes the lower
leaves are entire and alternately borne on the stem. The leaf lobes are lanceolate or
oblong, the mid-lobe is longest, serrate, glandular, on the midrib beneath, often
scorched with purple. Petioles are 1 ½ -2 ½ inch. long and reddish-purple. Stipules
are½ inch long, linear, acute. Flowers are borne singly in the axil upto 12.5 cm wide
yellow or buff with a rose or maroon eye. Pedicels are very short, stout, jointed near
the base and purple in colour. Involucral bracts are 10, lanceolate, shorter than the
calyx, adnate to its base, purple, Calyx glabrous becoming fleshy and bright red in
fruit (Sivarajan and Pradeep, 1996 and Singh, 2010), lobes lanceolate three nerved,

38
purple and together with the involucre acrescent, in fruit. Corolla is yellow with
purple at center. Stamens numerous, forming a column of 2 cm length, pink in colour;
ovary is superior and 5-celled, style with 5 branches. Roselle is a self pollinating plant
(McClintock and El-Tahir, 2004). But Vaidya, (1994) noticed natural cross
pollination in roselle. There is insect pollination. Abdel-Moniema et al. (2011)
reported insect prevailance on the roselle plants and their efficiency of pollination.
They reported that total of 16 species of insects belonging to seven orders (Hemiptera,
Homoptera, Lepidoptera, Herothera, Coleoptera, Diptera and Hymenoptera) are found
on the plants in two seasons. Honey bees are also found to be frequent visitors around
10 am to mid day. Fruit is ovoid, beaked, hairy and velvety. Capsules are 5 valved
with each valve containing 3-4 seeds. Seeds are large, black-brown, closely covered
with minute stout stellate hairs (Yadav and Sardesai, 2002 and Mahadevan et al.,
2009).
Floral formula:
3. Anatomy
Changdee et al. (2008) performed anatomical studies of tap roots in 10 days
old plants. Tetrarch xylem was noticed in the root. Casparian bands were found only
in the root basal part of roselle root exodermis and endodermis. Larger aerenchyma
like intercellular space was observed in cortex. Shaheen et al. (2009) reported
occurrence of eglandular twisted conical trichome (unicellular axillary elongated
broad at the base and tapering above) on both adxial and abaxial surfaces of roselle
leaf. Investigations of Dave et al. (1987) revealed that, roselle fruit pericarp has been
differentiated into epicarp, mesocrp and endocarp. Epicarp is having vacuolated,
compressed and tangential elongated cells with 5-7, layered thick mesopcarp and 3-5
layered endocarp. Presence of sclerides and fibres of diverse shapes at maturity was
noteworthy.
4. Cytogenetical Studies
Hibiscus sabdariffa is a tetraploid plant with n=18 and 4n=72 (Duke, 1983).
Ibrahim and Hussein, (2006) based on the study of 16 roselle genotypes concluded

39
that plant height, number of branches and dry sepal weight with high variability
should be taken in consideration during selection for higher yield of roselle. Tejaswini
et al. (1995) reported Heterosis, potency ratio and inbreeding depression in roselle (H.
sabdariffa (L.) var. altissima). Sobhan and Husain (1986) studied inheritance in
roselle; heterosis was observed for plant height, base diameter and length of fruit.
They observed monogenic inheritance for both length and diameter of fruits and that
involving 2.6 genes for most of the other characters. Plant height was also found to
vary markedly with seasonal variations while no variation was recorded among
varieties within years (Kumar et al., 1985). The additive gene action was indicated by
plant height while other characters exhibited both additive and multiplicative gene
actions. The characters revealed a wide variation in heritability, which ranged from
0.02 to 0.85 for number of epicalyx per fruit to base diameter of stem respectively.
Yield characters variability in roselle was studied by Atta et al. (2011). Seed yield
was found in the range of 292±8.80 kg ha-1 (E8) to 497±8.91 kg ha-1 (E4) while, calyx
yield varied from 123±8.26 kg ha-1 (E1) to 716±36.81 kg ha-1 (E9).
Mukewar et al. (1997) performed combining ability analysis in roselle. Their
di allele analysis showed predominance of additive gene action for seed yield per
plant and number of capsules per plant. Genotypes RIN-051, RIN-049 and RIN-096
exhibited positive significant GCA (General Combining Ability) effects for seed yield
per plant. Thirthamallappa and Sheriff (1992) also reported combining ability in
roselle.
Chakravarty and Basu (1972) studied anthocyanin pigmentation pattern in
roselle with special reference to the variation intermediacy and mode of inheritance.
They also reported that the difference in bushy and non-bushy habit and edible and
inedibleness of calyx are monofactorially and independently inheritable. Chakravarty
et al. (1981) studied new plant type in roselle which showed difference over other
types, in inheritance of two of its main distinguishing characteristics i.e. in glandless
(gs) sepal is recessive to normal glands (GS) and normal ovoid shape of capsule (Co)
is incompletely dominant over round shape (co). However, there was no any linkage
between gs and co. Adamson and O'Bryan (1981) studied inheritance of
photosensitivity in roselle. They found that in the food-type roselle, photo-sensitivity
is governed by two dominant genes both of which are required for expression of

40
character (duplicate recessive epistasis). However, in the fibre-type and wild-type
roselle, two dominant genes are involved and either will produce photosensitivity
(duplicate dominant epistasis).
Singh and Mitra, (1967) studied x-ray irradiation effects on roselle. There was
induction of mutation at high doses of X-rays. Shome, (1981) noticed that the 1kR to
2kR dose is optimal for inducing the mutations in seedling of roselle. Singh and
Tiwari, (1984) observed modification of mutagenic effectiveness and efficiency of
ethyl methanesulfonate (EMS) with streptomycin in roselle. Singh, (1988) reported
induction of mutations leading to mutants with seeds, superior in fats, fatty acids and
amino acid contents. Vaidya, (1994) was successful in developing induced female
sterile mutant in roselle.
Gassama-Dia et al. (2004) studied direct genetic transformation of roselle
using Agrobacterium LBA 4404 strain having pBal plasmid with β-glucuronidase
p35SGUSINT and plant selectable marker Neomycin Phospho-Transferase gene
(nptII). Their study revealed that 54.3% of the seedlings grew well on the selective
medium; 68% of the explants excised from putative transformed plants were found to
be GUS positive. After 60 days, evaluation by PCR showed nptII gene in roselle line.
This is perhaps the first report of genetic transformation in roselle.
5. Physiological Studies
a. Seed Germination
Kalavathi, (1996) noticed progressive loss of viability of roselle seeds with
increase in the storage period. According to her studies, the seeds when packed in
700-guage polyethylene bag, could maintain 73 percent germination even upto 15
months of storage. The better germination was achieved in roselle by giving alternate
temperatures of 25/30 0C (Ellis et al., 1985). Between the two methods, the use of 'top
of paper' or 'between paper' for conduct of germination, ‘between paper’ method is
best suited for the germination of roselle seeds (Kalavathi, 1996). Promolin (GA4 +
GA7 + Benzyladenine) at a concentration of 80 ppm was effective in improving the
germination of roselle seeds (Ynoue et al., 1996).

41
b. Growth
Sermsri et al. (1987) performed growth analysis of roselle (H. sabdariffa var.
altissima L.) in Thailand. With increasing age, there was rapid increase in total dry
weight up to 120 day stage and it decreased thereafter by complete stoppage at 150
days (2142 g m-2). Relative growth rate was found stable from 30-90 day after
planting. The ratio of each organ to total dry weight was found highest in leaf, which
was about 60 % during first few weeks and decreased with increasing age up to 6 % at
183 DAP. Stem showed increased ratio from 34 % to 84 % at 183 DAP. Leaf area
ratio (LAR cm2g-1) was decreased linearly from 86.8 to 10.6. However, leaf area
index (LAI m2 m-2) increased up to 6.3 at 120 DAP and decreased suddenly to 2.3 at
183 DAP. Thus, the count of net assimilation rate was about 8.8 g m-2 leaf day-1. Crop
growth rate (CGR) was found highest (34.94 g.m-2.day-1) in the period from 60-90
DAP. The relative growth rate (RGR) was highest, 13.2 % per day during first stage
and in later stage linear decrease was noticed. Atta et al. (2010 a) noticed that in three
ecotypes of roselle in Niger, LAI value ranged from 0.5 cm2cm2 to 1.0 cm2cm2
between 13 and 48 days after sowing. Thereafter LAI increased markedly for all
ecotypes with a maximum range between 62 and 83 days after sowing. The LAI
decreased to reach values between 1.0 cm2cm2 and 2.0 cm2cm2 three week later.
Analysis of Small and Rhoden (1991) revealed that stem diameter increased from 3.5
to 0.1 mm per week between 3rd and 9th week after transplantation. For calyx
production wide spacing (about 100 cm apart) is preferred while for fibre production a
close spacing of drilled seeds is essential i.e. 15 cm X 20 cm or 10 cm X 30 cm
(McClintock and El-Tahir, 2004). According to Jan (2010), row spacing (S) 60X80
and 80X80 cm had significantly affected the plant height, number of branches plant-1,
number of leaf plant-1, single fresh calyx weight and number of fresh calyx plant-1.
However, significant difference was not noticed in case of relative growth rate (RGR)
and number of fruits. Omer et al. (1997) noticed that pinching (45, 75 and 105 days)
is effective in increasing number of branches, weight of sepals and fruits, fixed oil and
anthocyanin in both dark and light coloured early maturing varieties of roselle.

42
c. Influence of Plant Growth Regulators on Growth, Development, Yield
and Chemical Composition
Influence of plant growth regulators on roselle has been studied by some
workers, Govinden-Soulange et al. (2009) studied effect of PGRs on rooting.
Softwood and semi hardwood cuttings from two-months-old plants were rooted on a
medium containing soil, compost and rocks and after dipping in IBA (indole-3-yl-
butyric acid) or NAA (a-naphthalene acetic acid) at 0-1.0 g l-1. The experiment
revealed that rooting was significantly (P< 0.05) enhanced by the type of cutting and
the concentration of auxin used. However, softwood cuttings responded more
positively to auxin treatment and lower levels (0.5g l-1) of auxin stimulated leaf and
root formation. Sadak (2005) reported that inhibition of IAA oxidase results in auxin
accumulation which is reflected in stimulatory growth and yield in roselle. Foliar
application of IAA (50 and 100 ppm) caused increase in plant height, stem girth, no of
leaves, leaf area ratio, shoot/root ratio and chlorophyll content in roselle plants
(Mukhtar, 2007). IAA treatment also led to increase in minerals (P, K, Mn and Fe
contents), carbohydrates and vitamin in the leaf tissue. He further noticed that foliar
application of GA (50 and 100 ppm) caused increase in plant height, leaf number, leaf
area ratio, shoot/root ratio and chlorophyll content. The level of carbohydrates,
proteins, vitamins and minerals (P, K, Cu and Zn) in the roselle leaf tissue was
increased due to GA application. Fathima and Balasubramanian, (2006) noticed
positive effect of plant growth regulators (GA 100+NAA 50 µgml-1) on the yield and
quality of bast fibres in H. sabdariffa L. var. altissima Webster.
Hassanein et al. (2005) found that foliar application of benzyladenine and/or
gibberellins (GA3) at 100 and 200ppm increased number of fruits/plant, fresh weight
of fruits/plant, fresh and dry weight of sepals/plant. The most effective dose was
100ppm GA + 100ppm BA in presence of Fe-EDTA. Foliar PGR treatments increased
anthocyanin and soluble sugar content in sepals and decreased titratable acidity. The
levels of mineral elements (K, Ca, Mg, P, and Fe) were increased due to BA and GA
treatment. The phenol metabolism was also promoted by foliar application of these
two PGR. Foliar application of Benzyladenine was found to promote nutrient
transport to the seeds and total nitrogen in seeds (Talaat and Youssef, 1998 and El-

43
Meleigy, 1989). El-Meleigy, (1989) further noticed that BA enhanced the
biosynthesis of anthocyanins in roselle leaves and calyx.
Mostafa et al. (2005) studied effect of foliar application of benzyladenine and
gibberellin (100 and 200 mg l-1) on seed yield and seed quality of roselle plants. They
found that foliar application of GA and BA increased the weight of seed/plant. The
spray of these PGRs significantly increased total soluble sugars, polysaccharides, total
carbohydrates, total soluble N, protein N, total N and oil content of seed of roselle.
The GLC analysis of roselle seeds revealed that palmitic acid was major among
saturated fatty acids while oleic acid and linoleic acids were major among unsaturated
fatty acids. However, there was decrease in palmitic acid due to GA3 and/or BA foliar
treatments in absence and presence of Fe-EDTA. Response of roselle to growth
retardants has been also investigated. Omer et al. (1997) reported that foliar
application of B-9 and CCC caused increase in oil yield in sepals of two early
maturing varieties of roselle.
d. Flowering and Photoperiodism
Mansour (1974) noticed that the roselle plant shows ambiphotoperiodic nature,
flowering both in short days and in extremely long days but remaining time it shows
vegetative nature (at least at 21-250C). He further reported that flowering had a dual
effect on growth. Strong floral induction caused lateral buds to develop into flowers
and the number of branches was proportionally reduced during first stage of
induction, apical dominance decreased. Plants shifted to long days after only a short
day at an inductive daylength and plants moved to short days after prolonged
placement in long days formed many more branches and much greater vegetative
mass than plants raised in continuous short days or long days. According to him
plants are sensitive to daylength even at cotyledonary stage. However, it is noticed
that roselle plants cannot be grown successively throughout the year as the species is a
typical short day plant (Morton, 1987). Roselle flowers best when the daylength is
shorter than 12 hrs and premature flowering is prevented at 13h daylength
(McClintock and El-Tahir, 2004).
e. Mineral Nutrition
Atta et al. (2010 b) studied changes in macroelement and micronutrients (Fe,
Mn, Cu, and Zn) contents at three different growth stages (I vegetative, II flowering

44
and III maturity) in leaves of three roselle ecotypes. In ecotype A7 nitrogen content
increased significantly between stages I and II then remaining constant up to stage III.
In ecotype A3 and A9 nitrogen content decreased significantly between stage II and
III. For all ecotypes leaf P content decreased progressively during plant growth. The
leaf Mg content was relatively constant in the three ecotypes at a given stage of plant
development. But it increased markedly from flowering to maturity. In the three
ecotypes Ca content remained constant from stage I to stage II then increased
considerably from stage II to III. Potassium content increased significantly in all the
three ecotypes from stage I to II. In all the three ecotypes, Fe and Zn contents
decreased significantly from stage I to stage II. The Mn content of all the three
ecotypes was similar at stage I thereafter it decreased continuously during plant
growth. In one ecotype (A9), the leaf Cu content was relatively constant during entire
growth cycle while, in other two ecotypes it decreased along with progress of growth.
These workers concluded that at 25 days growth stage maximum amount of these
micronutrients are present in leaves. Musa and Ogbadoyi, (2012) studied the effect of
nitrogen fertilizers on the levels of some nutrients and antinutritional and toxic
substances in roselle. They reported the presence of total oxalates, cyanides, nitrate
and some micronutrients viz. vit-C, β-carotene (precursor of vit.-A) and mineral
elements (Fe, Mg, Zn, Cu, Ca, Na and K) in roselle. Application of nitrogen fertilizers
significantly enhanced the levels of nitrate and β-carotene while, the level of vit.-C,
soluble and total oxalates in vegetable was decreased and other parameters were
unaffected. According to Nadia Gad (2011), application of cobalt at 20 ppm on
organically fertilized roselle plants was more effective in increasing growth, yield
quantity, anthocyanin, flavones and mineral contents (N, P, K , Fe, Mn, Zn and Cu).
Aziz et al. (2007) studied effect of cobalt and nickel on plant growth and flavonoid
content of roselle. They noticed that 20+2 mg kg-1 concentration of Co and Ni in soil
is beneficial for increasing plant height, no of branches per plant as well as fresh
weight and dry weight of plant. This concentration is quite effective in enhancing the
levels of mineral elements (N, P, K, Co, Ni, Mn, Zn and Cu) in leaves and calyx of
roselle plant. Also the highest contents of anthocyanin and flavones were noticed in
response to the same combination of Co and Ni.

45
f. Gas exchange and Photosynthesis
Ramachandra Reddy and Das, (1986) studied correlation between biomass
production and net photosynthetic rates and kinetic properties of RuBP carboxylase in
certain C3 plants including roselle. They noticed that the photosynthetic rates and
ribulose 1, 5-bisphosphatase carboxylase activities were well related to biomass
production. The superior biomass yields of roselle were positively correlated with
high photosynthetic capacity and low Km (CO2), as well as high Vmax values of
RuBP carboxylase in leaf. They concluded that the carboxylase efficiency and
photosynthetic performance are major determinants of biomass production in C3
plants in general and roselle in particular. According to Baker and Kassam (1975), the
dry matter in Roselle was lost due to dark respiration. About 91.32 % of loss of dry
weight was noticed in 24 h period.
g. Stress Physiology
i. Water stress
Salama et al. (2001) studied effect of decreasing soil water potentials on
chlorophyll pigments in roselle at various osmotic potentials. The amount of
chlorophyll was decreased with increasing osmotic stress from -3 to -13 bars. The
foliar treatment of kinetin increased Chl-a contents in unstressed as well in stressed
plants. However, Chl-b was increased only under stress conditions. Chl-a/Chl-b ratio
was not significantly increased under stress conditions. Chl-a/Chl-b ratio was
increased at -3 and -10 bars soil water potential but not at -13 bars. The chlorophyll
stability was also checked in which Chl-a and Chl-b was significantly increased with
decreasing soil water potentials, only exception was -7 bars. The spray of kinetin
significantly enhanced Chl-a stability except unstressed plants at -13 bars. Chl-a and
Chl-b stability were significantly increased with increasing soil water matric potential,
except only at -3 bars. The kinetin treatment significantly enhanced Chl-a stability
under stress conditions.
ii. Waterlogging stress
Changdee et al. (2008, 2009 and 2010) studied effect of waterlogging on the
growth and root development as well as some yield parameters. They noticed a
marked effect of prolonged waterlogging on various growth characteristics such as
plant height, stem diameter, leaf area biomass production. Under waterlogging

46
conditions there was decline in plant height and fibre yield (Changdee et al., 2010).
Biomass production was also affected due to waterlogging. Roselle subjected to 30,
60, 90 and 120 DAS had reduced plant height by 26.2, 23.3, 7.7 and 1.2 %
respectively. Stem diameter was reduced to some extent at 120 DAS (2.2 %). Effect
of waterlogging on leaf area of roselle was prominent i.e. 5.4 % in treatments at 120
DAS (Changdee et al., 2010). Above ground, biomass was also affected due to
waterlogging. In roselle leaf dry weight, wood dry weight, bark dry weight and shoot
dry weight was reduced significantly in all treatments over control (Changdee et al.,
2010). It had its equal negative influence on bark thickness, fibre yield, root dry
weight of roselle. Changdee et al. (2009 and 2010) noticed a higher reduction in fibre
yield under waterlogging conditions in roselle. However, increased adventitious root
growth formation took place under waterlogging conditions. These adventitious roots
were with well developed aerenchyma (Changdee et al., 2008). However, onset of
endodermal casparian bands appeared slightly closer to root tips as compared to
control (Changdee et al., 2008). Root anatomical studies were carried out in roselle
subjected to waterlogging conditions and it was noticed that the formation of
exodermal casparian bands occurred only at the basal portion of tap roots. According
to Changdee et al. (2008), this plant species exhibits flooding tolerance to some extent
and may be recommended for flooding prone-environments.
iii. Salinity
Simoens and Van Damme (1988) compared the effects of soil salinity on the
germination capacity of two Senegalese varieties of roselle. The germination
percentage remained high in the 0-9 mScm-1 range for the red variety (H. sabdariffa
L. var. altissima type Red) and in the 0-5 mScm-1 range for green variety (H.
sabdariffa L. var. albus). Based on germination studies and performance of mature
plants they concluded that roselle is salt sensitive and green variety was more
sensitive than red variety. Bijeh keshavarzi and Moussavinik (2011) studied seed
germination, seedling growth (radicle length, plumule length, seed vigour, fresh
weight and dry weight) under different NaCl salinity regimes in roselle. They noticed
decline in all the studied parameters at 150 mM salinity. El-Saidi and Hawash (1971)
studied the effect of saline water irrigation on the growth and chemical properties of
roselle plants. El-Sharkawi (1986 a, b) reported that increase in sodicity reduced

47
soluble proteins and total free amino acids in this species. These workers further
noticed that chlorophyll content was not sensitive to sodicity.
Ali et al. (2012 a) studied the effects of gibberellic acid on growth and
photosynthetic pigments of roselle growing under saline conditions. They reported
that GA3 application reduced the inhibitory effects of NaCl salinity on growth and
photosynthetic pigments by inducing the enzyme carbonic anhydrase (CA) activity
and enhanced relative water content. In this way GA3 plays important role in salt
stress tolerance in roselle plant. Abdel Latef et al. (2009) noticed positive effect of
benzyladenine (BA) on saline injury in two roselle cultivars via equilibrium of
cytosolutes including anthocyanins. Their study revealed that 250 mg l-1 spray of BA
could partly alleviate the salt stress symptoms in both cultivars.
iv. Low temperature stress
Roselle can satisfactorily grow between 18 0C to 35 0C. McClintock and El-
Tahir (2004) have mentioned that the plant growth stops at 14 0C and death occurs
after 15 days. Exposure to 10 0C for 2-3 days also causes death of the plants and
cotyledons cannot withstand 2-3 h exposure below 10 0C. Flower and calyx
production is affected due to temperature below 17 0C. These observations clearly
indicate that roselle is highly sensitive to low temperature stress.
6. Tissue culture studies
Sie et al. (2010) described in detail protocols for callus and somatic embryo
initiation in roselle and also explained influence of explants types, sugar and plant
growth regulators on the process. They tested two genotypes (H. sabdariffa var.
sabdariffa and H. sabdariffa var. altissima), two sugars (sucrose and glucose) and
three concentrations (1 %, 2 % and 3 %) of each sugar. Root, hypocotyl, cotyledon
were used as explants for tissue culture. Fourteen combinations of plant growth
regulators (PGRs) in MS medium and five combinations of PGR in Driver and
Kuniyuki (DKW) medium were tested on hypocotyl and cotyledon for callus and
somatic embryo formation. The PGR combinations used with MS medium were
naphthaleneacetic acid/kinetin (NAA/KIN), 2, 4-dichlorophenoxyacetic acid/kinetin
(2, 4-D/KIN), and naphthaleneacetic acid/6-benzylaminopurine (NAA/BA) and those

48
used with DKW medium were 2,4-dichlorophenoxyacetic acid/thidiazuron (2,4-
D/TDZ). All concentrations of both sugars and PGRs were found efficient in
successful induction of callus in all explant types of both genotypes. The best results
for callus induction were achieved with 3 % sucrose in the hypocotyl and cotyledon
explants. Somatic embryos were obtained with DKW medium supplemented with 4
mg l-1 2, 4-D + 1 mg l-1 TDZ and 1 mg l-1 2, 4-D + 0, 5 mg l-1 TDZ. Sadia (2000) was
able to regenerate plants of roselle through axillary and branching under in vitro
conditions. The use of shoot apices to micropropagate roselle has been described by
Gomez-Leyva et al. (2008). Callus formation was observed on both TDZ and 2, 4-D
on leaf explants of roselle. However, 1.5 mg l-1 TDZ showed a higher response in
callus formation (80 %); nevertheless the lowest callus formation was observed with
0.1 mg l-1 of TDZ. On the other hand, all levels of 2, 4-D (0.01-0.05mg l-1) produced
equally high percentage of callus (80 %). Conversely, these calli could not be
regenerated further as only profuse chlorophyll formation (organogenic calli) was
observed after two weeks culture on different levels of KIN (0.1-5.0 mg l-1).
Govinden-Soulange et al. (2009) reported successful in vitro regeneration from nodal
explants of germinated seedlings, stem and leaf tissues of roselle. According to them,
in vitro regeneration seemed to be more successful by axillary bud regeneration than
by indirect organogenesis. As such, more efficient shoot regeneration from axillary
buds was obtained on MS media with low levels of cytokinins than on media
supplemented with higher concentrations of BAP or KIN. The optimum concentration
of cytokinin for successful shoot regeneration was between 0-0.1mg l-1for both
growth regulators. Gomez-Leyva et al. (2008) have described multiple shoot
generation in roselle through shoot apex (meristem) culture. Mizukami et al. (1988)
observed that from among secondary metabolites only cyanidin could be produced
through callus culture in this species. Mizukami et al. (loc. cit.) detected the activity
of enzyme flavones-3-hydroxylase which catalyze the conversion of naringenin into
dihydroflavonol in cultured cells of roselle. According to these workers this enzyme
may not be involved in regulation of anthocyanin biosynthesis in roselle. Mizukami et
al. (loc. cit.) evaluated time course changes in activity of anthocyanin accumulation
and activities of phenylalanine ammonia lyase and chalcone synthase in roselle callus
tissues incubated under different culture conditions. The activity of phenylalanine

49
ammonia lyase was not affected by light regime as well as supplement of either IAA
or 2, 4-D. On the other hand, chalcone synthase activity was markedly suppressed
when the callus was cultured in a medium containing IAA instead of 2, 4-D or in
darkness. These findings indicate that chalcone synthase plays more important role in
anthocyanin biosynthesis regulated by 2, 4–D and light irradiation than phenylalanine
ammonia lyase.
7. Phytochemical constituents
The chemical composition of different parts of roselle has been studied by
number of workers. The composition of leaf is summarized in Table 9.
Table 9. Chemical composition of Hibiscus sabdariffa leaf.
Sr. No.
Component Quantity (%)
1. Moisture percentage 86.2
2. Protein content 1.7 3. Fat 1.1 4. Carbohydrate 10.0 5. Mineral matter 1.0 6. Ca 0.18 7. P 0.04 8. Fe 0.0054 9. Acid content 1.25
Source: Leung et al.(1968).
Ibnusaud and coworkers were granted US Patent (US 6127553 A) in the year
2000 for developing a process for isolation of Hibiscus acid ((+) hydroxyl citric acid
lactone) from leaves of H. sabdariffa L.
Calyx is perhaps the most thoroughly investigated plant part with respect to
phytochemistry. A proximate chemical composition of roselle fleshy calyx is
summarized below.
Table 10. Chemical composition of Hibiscus sabdariffa fleshy calyx.
Sr. No.
Component Quantity (%)
1. Moisture percentage
88.26

50
2. Crude protein content
1.46
3. Ether extract 1.97 4. Carbohydrate 5.86 5. Reducing Sugars 0.82 6. Sucrose 0.24 7. Crude fibre 1.58 8. Ash content 0.87 10. Ca 0.108 11. P 0.052 12. Fe 0.021
13. Mn, Al, Mg, Na and K
In trace
14. Acids Citric acid 3.74
Malic acid, Tartaric acid, Hibiscus acid (δ-lacton Hydroxy citric acid M.P. 181 B.P. 830C C5H6O7)
-
15. Pectin 3.19 Source: Webster, (1920), Health Bulletin, (1951), Sastri, (1959) and Leung et al. (1968).
Babalola et al. (2000) reported that the leaves and calyx of green roselle
variety are very rich in β-carotene, vit.-C and riboflavin with some mineral elements.
They also reported that ascorbic acid and crude fibre content of green coloured calyx
were significantly higher than the red and dark red roselle. Major organic acids
present in flower extracts were citric acid, malic acid and ascorbic acid (Buogo and
Picchinenna, 1937 and Reaubourg and Monceaux, 1940). Hida et al. (2005) stated
that hydroxycitric acid is a major acid present in calyx of H. sabdariffa. Kafaga and
Koch, (1980 a, b, c and d) studied effect of stage of maturity on quality of roselle with
respect to anthocyanins content from flowering to overripeness. The strains from
Central America and Thiland needed 45-50 days to reach this stage of maturity. They
noticed presence of organic acids such as citric acid, Hibiscus acid, malic acid and
tartaric acid, anthocyanins, mucilage pectin and carbohydrates in the calyces from
five strains of roselle varieties. Wong et al. (2002) studied physico-chemical
characteristics of roselle. They reported 141.09 mg 100 g-1 of ascorbic acid, 1.88 mg
100g-1 of β-carotene and 164.34 µg 100g-1 of lycopene. Mueller and Franz, (1992)
studied the chemical structure and biological activity of polysaccharides from roselle.
They reported that floral bud extracts have three water soluble polysaccharides HIB 1

51
and 2 composed of arabinans and arabinogalactans. Thin layer chromatography and
HPLC techniques were used to determine and identify anthocyanins. Delphinidin-3-
sambubioside and cyanidin-3-sambubioside are two main anthocyanins in roselle
(Wong et al., 2002 and Juliani et al., 2009). Rodriguez-Medina et al. (2009) reported
presence of phenolic fractions and polar compounds in calyx. Using HPLC technique
Segura-Carretero et al. (2008) reported presence of anthocyanins and chlorogenic acid
in dried calyces of roselle. They identified Delphinidin-3-Sambubioside, Cyanidin-3-
Sambubioside along with cyanidine-3-O rutinoside, delphinidin-3-O gluoside, and
cynidine-3-5-diglucoside and chlorogenic acid as minor constituents in aqueous
extract. The antioxidant compounds such as protocatechuic acid are also detected in
roselle calyx (Osman et al., 1975 and Tee et al., 2002).
The chemical analysis of roselle seeds has been carried out by some workers
(Samy, 1980, Rao, 1996, Hainida et al., 2008 and Nzikou et al., 2011). The proximate
analysis of the seed is given in Table 11.
Table 11: Proximate analysis of roselle seeds
Sr. No.
Component Reported values
1. Moisture Content (%) 5.57-9.93 2. Crude protein (%) 25.20-33.45 3. Ether extract (%) 21.10-22.13 4. Crude fibre (%) 16.30-18.3 5. Ash content (%) 5.19-7.47 6. Total carbohydrate (%) 21.25-26.64 7. Calcium (mg100g-1) 647 8. Magnesium (mg100g-1) 1329 9. Potassium (mg100g-1) 1329 10. Sodium (mg100g-1) 659 11. Phosphorus (mg100g-1) 510 12. Fe and Zn Highest 13. Total digestible nutrients (%) 75.8 14. Starch (%) 84.06 15. Amino acids (mg kg-1) 250-625 16. Tannin (mg kg-1) 7.46 17. Calorific value (Kcal 100g-1) 326.53
Source: Al-Wandawi et al. (1984); Ismail et al.(2008); Hainida et al. (2008) and Nzikou et al. (2011).
Seed proteins of different varieties of roselle have been analyzed for amino
acid makeup (Morton, 1987, El-Adawy and Khalil, 1994 and Rao, 1996). Some of
these findings are recorded in Table 12.

52
Table 12. Amino acid composition of seed proteins in different roselle varieties
Sr. No.
Amino acid Morton, 1987 El-Adawy and Khalil, 1994 Rao, 1996
Light red Early dark red
Late dark red
AMV-2 Bhimili-1
1. Aspartic acid 16.3 10.91 10.50 10.52 9.1 8.3
2. Threonine* 3.0 4.86 4.74 4.39 4.2 3.8
3. Serine 3.5 4.40 4.65 4.70 5.2 5.2
4. Glutamic acid 7.2 21.30 21.78 21.38 21.6 21.8
5. Proline 5.6 4.14 4.13 4.21 5.2 4.6
6. Glycine 3.8 4.27 4.27 4.32 6.2 5.1
7. Alanine 3.7 4.69 4.55 4.72 6.1 4.2
8. Cystine * 1.3 2.64 2.91 2.88 1.0 4.2
9. Valine* 3.8 3.26 3.32 3.32 5.0 4.8
10. Methionine* 1.0 1.13 1.20 1.15 1.2 1.9
11. Isoleucine* 3.0 3.24 3.03 3.64 5.0 3.7
12. Leucine* 5.0 7.32 7.29 7.28 6.5 5.9
13. Tyrosine* 2.2 3.46 3.39 3.54 2.7 3.2
14. Phenylalanine* 3.2 5.09 5.25 5.15 4.8 4.7 15. Lysine* 3.9 5.37 5.46 5.56 4.3 3.3 16. Histidine 1.5 2.97 2.74 2.77 NA NA
17. Tryptophan* - 0.37 0.29 0.34 1.2 1.4
18. Arginine 3.6 10.58 10.50 10.13 9.9 10.2 Source: Morton, (1987); El-Adawy and Khalil, (1994) and Rao, (1996). Note: *Essential amino acids according to FAO/WHO (1991). Values depicted in table are in percent.
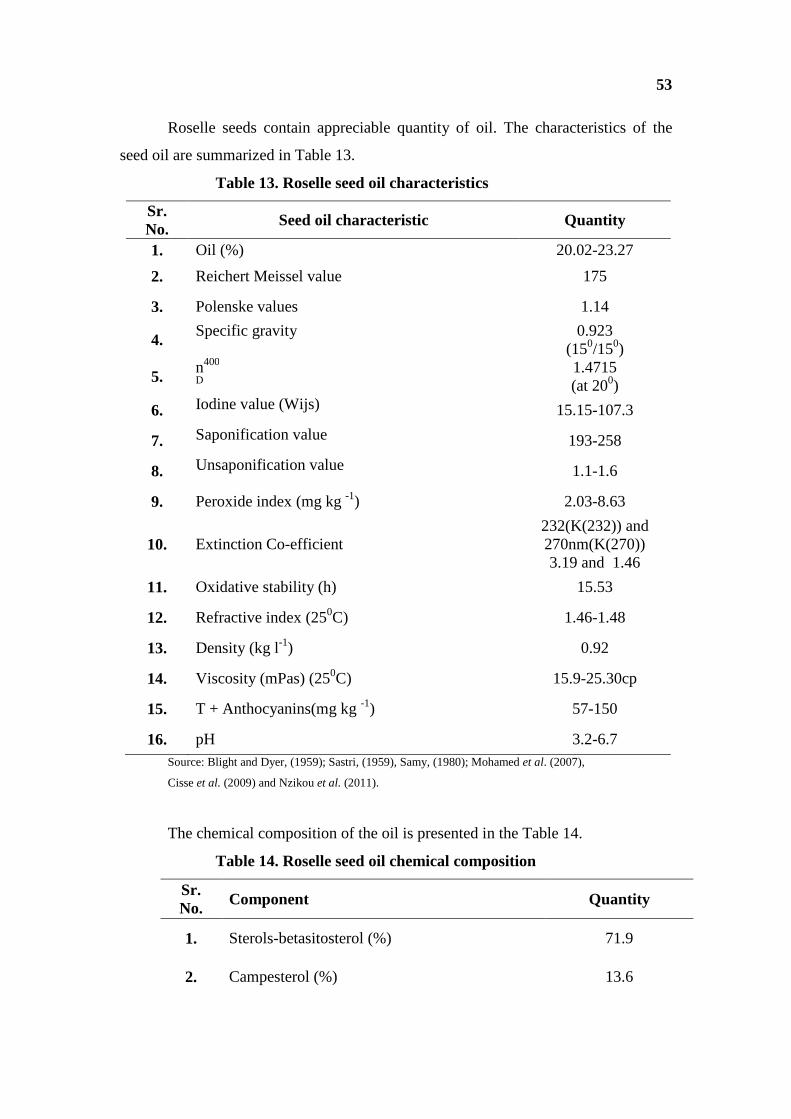
53
Roselle seeds contain appreciable quantity of oil. The characteristics of the
seed oil are summarized in Table 13.
Table 13. Roselle seed oil characteristics
Sr. No.
Seed oil characteristic Quantity
1. Oil (%) 20.02-23.27
2. Reichert Meissel value 175
3. Polenske values 1.14
4. Specific gravity 0.923
(150/150)
5. n400
D 1.4715 (at 200)
6. Iodine value (Wijs) 15.15-107.3
7. Saponification value 193-258
8. Unsaponification value 1.1-1.6
9. Peroxide index (mg kg -1) 2.03-8.63
10. Extinction Co-efficient 232(K(232)) and 270nm(K(270)) 3.19 and 1.46
11. Oxidative stability (h) 15.53
12. Refractive index (250C) 1.46-1.48
13. Density (kg l-1) 0.92
14. Viscosity (mPas) (250C) 15.9-25.30cp
15. T + Anthocyanins(mg kg -1) 57-150
16. pH 3.2-6.7 Source: Blight and Dyer, (1959); Sastri, (1959), Samy, (1980); Mohamed et al. (2007),
Cisse et al. (2009) and Nzikou et al. (2011).
The chemical composition of the oil is presented in the Table 14.
Table 14. Roselle seed oil chemical composition
Sr. No. Component Quantity
1. Sterols-betasitosterol (%) 71.9
2. Campesterol (%) 13.6

54
3. Delta-5-avenasterol(%) 5.9
4. Cholesterol(%) 1.35
5. Clerosterol(%) 0.6
6. Total-tocopherols (mg kg-1) 2000
7. α-tocopherol (%) 25
8. γ-tocopherol(%) 74.5
9. δ-tocopherol (%) 0.5
10. Acidity(%) 2.24-4.4
11. Fatty acids(%) 17
Source: Blight and Dyer (1959); Sastri, (1959); Samy (1980); Mohamed et al. (2007), Cisse et al. (2009) and Nzikou et al. (2011).
The results of fatty acid analysis of the roselle oil are depicted in Table 15.
Table 15. Fatty acid composition of Hibiscus sabdariffa seed oil
Sr. No.
Fatty acid Quantity (%)
A. Saturated acid 26.57-27.43
B. Unsaturated acid 72.57-73.4
1. Myristic acid (C14:0) 0.26-0.31
2. Palmitic acid (C16:0) 20-21.65
3. Palmitoleic acid (C 16:1) 0.44
4. Stearic acid (C 18:0) 5.3-5.97
5. Olieic acid (C 18:1) 28-32.28
6. Linoleic acid (C 18:2) 39.16-40
7. Linolenic acid (C 18:3) 0.57-1.69
8. (C 19:0) 1.7
9. Arachidic acid (C 20:0) 0.72

55
10. Eicosenoic acid (C 20:1) 0.08
11. Eicosatrienoic acid (C 20:3) 0.34
12. Behenic acid (C 22:0) 0.37
Source: Blight and Dyer, (1959); Sastri, (1959), Samy, (1980); Mohamed et al. (2007),
Cisse et al. (2009) and Nzikou et al. (2011).
Jirovetz et al. (1992) studied seed oil of roselle for the evaluation of the
volatiles by means of GC MS and GC FTIR. They reported presence of volatiles,
mainly unsaturated (one or two double bonds) hydrocarbons, alcohols and aldehydes
ranging from C8 to C13. Fang et al. (2004) performed isolation and structural analysis
of saponin from seed of roselle. A study done by El-Adawy and Khalil, (1994)
revealed that there are no distinct differences in three cultivars of roselle seeds from
Egypt and these showed a low tannin and phytic acid content and the absence of
hemagglutinin activity. Al-Wandawi et al. (1984) and Abu-Tarboush and Ahmed,
(1996) reported that whole roselle seeds had a lower percentage or trace of free and
bound gossypol compounds. Trypsin and α-chymotrypsin inhibitor activities in
defatted roselle seed flour were found to be lower than soybean defatted flour (Abu-
Tarboush and Ahmed, 1996).
8. Cultivation
Roselle a leafy vegetable which yields leaves and calyces is grown in home
garden and also cultivated on large scale in some regions. It is also grown as an
intercrop. It is considered as a low input and less labour demanding crop, which needs
less care with respect to weed control. Roselle can grow well in tropical as well as
subtropical regions at about 3000 ft. (900 m) up from sea levels. The crop is best
cultivated between 100N to 300S (Singh, 2010). In temperate regions it can be grown
as summer crop. According to Duke (1983), it can tolerate flood conditions and heavy
wind while, it is not shade loving as well as it does not withstand frost conditions.
However, it is a better drought resistant type (Singh, 2010). According to Ismail et al.
(2008) roselle requires a permeable soil preferably a friable sandy loam with humus
but it can adapt to variety of soils. Singh, (2010) has mentioned that soil pH 4.4 to 7.8
is suitable for roselle and higher soil pH causes chlorosis. The crop is generally sown
at the beginning of mid rainy season and mid April in India. Under low rainfall

56
conditions irrigation is advantageous. A well distributed rainfall of 1500-2000 mm
year-1 is optimum for roselle growth. The seeds are sown at a depth of 0.5 cm but 2.5
to 3 cm depth show better results for germination of species (Duke, 1983 and Singh,
2010). For broadcasting sowing, seeds rate is about 13-15 kg ha-1 and for line sowing
it is about 11-13 kg ha-1 (Morton, 1987). The seedlings after full germination are
thinned to keep 1m distance between two plants. Besides this usual method of
cultivation, propagation through rooted shoot cuttings is also practiced in some parts
of India (Mahadevan et al., 2009). Due to propagation of stem cuttings, shorter plants
are produced which are suitable for intercropping with tree crops but the calyx yield
of such plants is rather low. Weeding for the first month is needed with a
simultaneous thinning of seedling. Generally harvesting is done with the help of
sickle for the fibre purpose after 3-4 months and sometimes after 10 months in
Indonesia, while, for the calyces of fruits, about 3 weeks after the onset of first fruits
are ready for picking (Duke, 1983). The fruits of roselle ripen progressively from the
lowest to the highest and harvesting of fruit is done when these are fully ripened
(Mahadevan et al., 2009). Calyx production ranges from a 1.5 kg ha-1(calif) to 2 kg
ha-1 (Puerto Rico) to 7.5 kg ha-1in South Florida, 16000 kg ha-1in intercropping, 19000
kg when planted alone. Dual purpose planting can yield 17000 kg ha-1 of herbage in 3
cutting and later 6300 kg ha-1of calyces (Morton, 1987).
According to Singh (2010), there are many promising roselle varieties
cultivated in India. The information about these cultivars is briefly summarized in
Table 16.

57
Table 16. Roselle varieties and their life style
Sr. No. Variety
Research Station
Selection/ Hybridizati
on Stem Leaf Flowering period Seeds
Sowing period and yield
1.
HS-4288
CRIJAF Barrakpore
RT-1 (non bristled)X RT-2 (bristled)
Green with Red pigment at the node
Petiole upper surface is green Lower surface is red
180 days Coffee coloured and reniform
Mid April to May Yield: 18-20 q ha-1 Area: West Bengal, Assam, Tripura and Bihar.
2. HS-7910
CRIJAF Barrakpore
HS-4288 X RT-1
Green coloured With flash of red pigment
Deeply lobed, more reddish,
180 days Coffee coloured, reniform
May, Yield: 18-20 q ha-1 Area:WestBengal, Assam, Orissa and Bihar.
3. AMV-1 Agriculture Research Station Amadalavalasa (A.P.)
It is the result of selection made from an indigenous material.
Crimson purple, Bristled
Deeply lobed, purple petiole.
180-200 days Brown in colour
Mid April to May Yield: 15-17 q ha-1 Area: A.P. and Orissa.
4. AMV-2 Agriculture Research Station Amadalavalasa (A.P.)
It is the result of the selection made from an indigenously collected material.
Pink coloured Fully bristled
Deeply lobed 180-200 days Brown in colour
Early May Yield: 17 q ha-1 Area: A.P. and Orissa
5. AMV-3 Agriculture Research Station Amadala
AMV-1 X ER-79
Pink coloured with bristles
Deeply lobed 180-200 days brown in colour
Early May Yield:18 q ha-1 Area: A.P. and Orissa
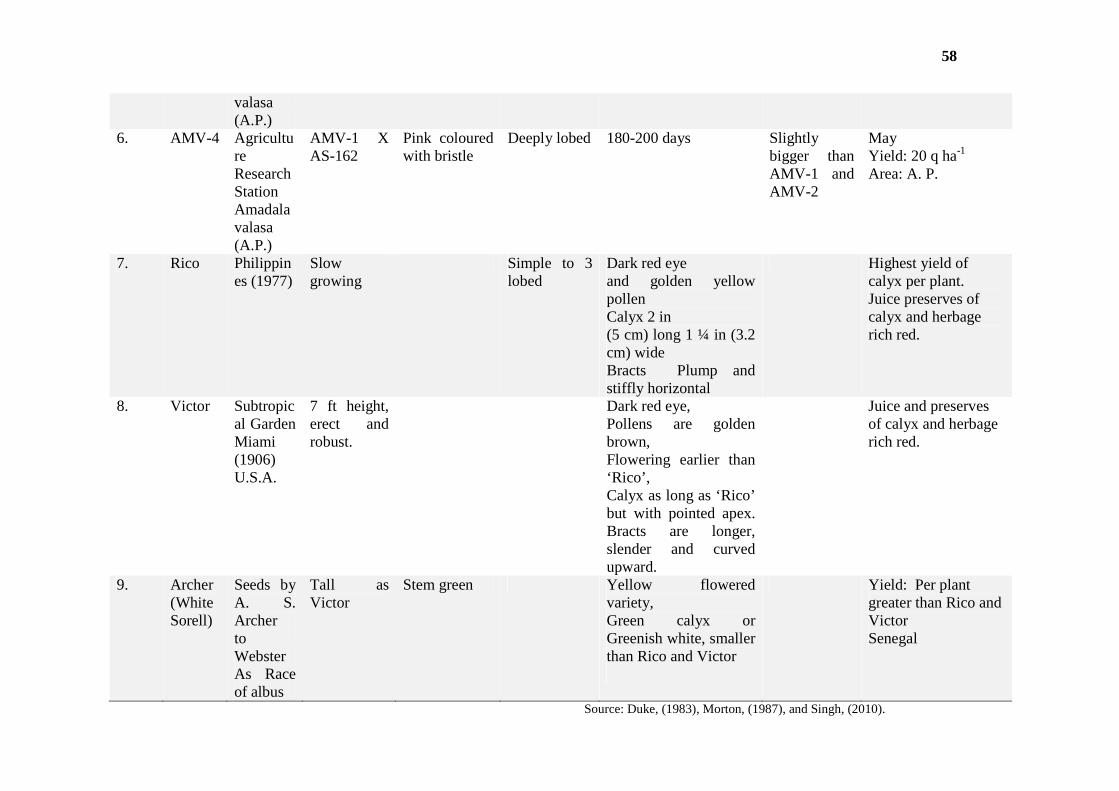
58
valasa (A.P.)
6. AMV-4 Agriculture Research Station Amadalavalasa (A.P.)
AMV-1 X AS-162
Pink coloured with bristle
Deeply lobed 180-200 days Slightly bigger than AMV-1 and AMV-2
May Yield: 20 q ha-1 Area: A. P.
7. Rico Philippines (1977)
Slow growing
Simple to 3 lobed
Dark red eye and golden yellow pollen Calyx 2 in (5 cm) long 1 ¼ in (3.2 cm) wide Bracts Plump and stiffly horizontal
Highest yield of calyx per plant. Juice preserves of calyx and herbage rich red.
8. Victor Subtropical Garden Miami (1906) U.S.A.
7 ft height, erect and robust.
Dark red eye, Pollens are golden brown, Flowering earlier than ‘Rico’, Calyx as long as ‘Rico’ but with pointed apex. Bracts are longer, slender and curved upward.
Juice and preserves of calyx and herbage rich red.
9. Archer (White Sorell)
Seeds by A. S. Archer to Webster As Race of albus
Tall as Victor
Stem green Yellow flowered variety, Green calyx or Greenish white, smaller than Rico and Victor
Yield: Per plant greater than Rico and Victor Senegal
Source: Duke, (1983), Morton, (1987), and Singh, (2010).

59
a. Fertilizer Application
Roselle gives good response to fertilizers. A positive response of calyx yield,
pod/plant and seeds/pod to nitrogen fertilizers was recorded by Oyewole and Mera,
(2010). Small and Rhoden, (1991) noticed that nitrogen application (100 and 200 mg
l-1 irrigation water) caused increase in dry matter, plant height and stem diameter. But
at the same time, Atta et al. (2010 a) have mentioned that with the exception of
number of branches/plant, yield components were not influenced by nitrogen
application and 50 kg N ha-1 may be considered as the optimum fertilization for leaf
yield. Lakshminarayana et al. (1980) studied the efficiency of nitrogen utilization by
Roselle. The field trials conducted by them (1975 and 1976) with Roselle cultivar
AMV-1 and AS7 revealed that fibre yield increased with increase in N application
from 0 to 80 kg N ha-1 and AMV-1 gave the highest yield as 1.09 t ha-1.
Commercial fertilizer application is also found effective with a stable manure
dose. Plant growth and sepal yield were considerably enhanced due to the application
of Nitrogen fertilizers in combination with chemical fertilizers application and
biofertilizers or indirectly through microbial propagation (Shaalan et al., 2001 and
Hassan, 2009). Plant height and number of branches per plant were significantly
increased due to application of nitrogen fertilizers because of importance and
involvement of nitrogen in vegetative growth of roselle plant (Okosun et al., 2006 and
Atta et al., 2010 a). Several workers also noticed similar response to NPK fertilizers
in roselle plant (Okosun, 2000, Babatunde, 2001, Babatunde et al., 2002 and Okosun
et al., 2006). The N, P, K fertilizers were effective at 2 g l-1 rate than lower rates. The
red cultivar gave best results for all the parameters. Abbas and Ali (2011) studied
effect of foliar application of NPK at the rate of 0, 1 and 2 g l-1 on some growth
characters of two cultivars of roselle, red and lined. Plant height, number of branches
per plant, total number of fruits and dry calyces yield were increased after foliar
application of NPK. The growth promotion also took place in terms of increase in
chemical constituents (vit-C, titratable acidity and total anthocyanin) due to
application of NPK fertilizers (Abbas and Ali, 2011). Increase in carbohydrate content
and photosynthetic pigments due to foliar application of NPK fertilizers, was also
evident (Abbas and Ali, 2011). Akanbi et al. (2009) reported that combination of
organic manure (5 ton ha-1compost) and inorganic fertilizer (NPK, 150 kg ha-1) is

60
optimum for good growth and high quality roselle production. The crop gave good
response to fertilizer application at the rate of 80 kg N ha-1 36-54 kg P2O5 ha-1and 75-
100 kg K2O ha-1 and 153 kg ha-1 for better growth in Java (Duke, 1983 and Singh,
2010). Abo-Baker and Mostafa, (2011) noticed that seed inoculation with
Azospirillium species and Bacillus polymyxa combined with 50 % chemical fertilizer
improved growth characters, yield components and chemical constituent of roselle
plants. Hassan, (2009) also recorded that by applying biofertilizers alone or in
combination with chemical fertilizers there was significant improvement in the
chemical constituents (total anthocyanin, total carbohydrates and chlorophyll
contents).
b. Diseases and Pests
Several pathogens and pests are found to attack plants of roselle (Duke, 1983
and Morton, 1987)
ü Fungal Diseases: Roselle is infected by several fungi. Aecidium
garckeanum, A. hibiscisurattense, Alternaria macrospora,
Cercospora abelmoschi, C. malaysensis, Corynespora
cassiicola, Cylindrocladium scoparium, Diplodia hibiscina,
Fusarium decemcellulare, F. sarcochroum, F. solani, F.
vasinfectum, Guignardia hibisci-sabdariffae, Irenopsis
molleriana, Leveillula taurica, Microsphaera euphorbiae,
Phoma sabdariffae, Phymatotrichum omnivorum,
Phytophthora parasitica, Phytophthora terretris, Pythium
perniciosum, Rhizocotonia solani, Sclerotinia fuckeliana, S.
sclerotiorum, Sclerotium rolfsii are some fungi attacking roselle
plant. Najafinia et al. (2004) reported occurrence of root and
crown rot caused by Cylindrocarpon didymium.Agbenin et al.
(2006) reported Fusarium wilt on red calyx roselle in Northern
Nigeria. According to Singh (2010), the most serious diseases
of roselle in India are leaf spot caused by Phoma sabdariffa,
fruit and stem rot caused by Phytophthora parasitica var.
sabdariffae.

61
ü Viral Diseases: Viral diseases such as Leaf curl (Abbaiah et
al.,1997), Cotton leaf curl and Yellow vein mosaic were
reported on roselle. Chatterjee et al. (2005) noticed that roselle
plants are attacked by Begomovirus leading to yellow vein
mosaic disease.
ü Bacterial Diseases: Roselle plants also suffer from few
bacterial diseases. Maconellicoccus hirsutus infection and its
control have been reported by Das and Singh, (1986). Amusa,
et al. (2001) noticed that roselle plant suffered by stem and
foliar blight in the tropical forest region of South Western
Nigeria.
ü Root-knot disease: Root knot nematodes like Meloidogyne
arenaria, M. incognita acrita, Heterodera rudicicola. and M.
javanica are reported on roselle plants (Duke, 1983; Morton,
1987; Agbenin and Ogunlana 2006 and Singh, 2010).
ü Insect pests: Major insect pests attacking Roselle plant are
Anomis erosa, Chaetocnema spp., Cosmophila erosa,
Dysdercus cingulatus, D. poecilus, Drosicha townsendi,
Nistora gemella, Phenacoccus hirsutus, Pseudococcus
filamentosus and Tectocoris diophthalmus. In Australia, 3
beetles, Nisotra breweri, Lagris cyanea, and Rhyparida
discopunctulata, feed on the leaves.
9. Uses
a. Human Nutrition
i. Food
The fruits of roselle are edible (Watt and Breyer-Brandwijk, 1962). According
to Tanaka (1976), tender leaves and stalks of this species are eaten as salad and used
for seasoning curries. He has further mentioned that dried calyces are used as potherb.
The leaves and fresh green calyces are used to make soup in Africa while in USA the
leaves and young shoots are also eaten raw in salads and red fleshy calyx lobes are
chopped and used in fruit salads (McClintock and El-Tahir, 2004). For preparation of

62
vegetables, green roselle types are preferred over red types. Fleshy roselle calyces are
used in wine jelly, syrup, gelatin, refreshing beverages, jams, sauces, pudding and
cakes. Dried roselle is used for tea, jelly, marmalade, ices, ice-cream, sherbets, butter,
pies, sauces, tarts and other desserts outside India. In India fleshy fruiting calyces are
used in preparation of sauce or jelly (Sivarajan and Pradeep, 1996). Seeds of roselle
are roasted and eaten. Ismail et al. (2008) have reported that in Africa, the seeds are
roasted and ground into a powder then used in oil, soup and sauces as a meal for
human consumption. According to Rao (1996), the cooked Mesta seeds can be
consumed as good quality food, with proteins rather than raw consumption. Yagoub et
al.(2004) stated that ‘Furunda’ a meat substitute is traditionally made by cooking
seeds and fermenting them for 9 days. Roselle seeds are also used to prepare
‘Bikalga’ an alkaline fermented food in Africa (Ouoba et al., 2009). The oil of roselle
seed is used for cooking in Chad, Tanzania and China (McClintock and El-Tahir,
2004).
ii. Beverages
Among different plant parts of roselle, calyces are perhaps the most popular
and commercially most exploited. The fresh as well as dried calyces have been
extensively used for preparation of beverages. These are consumed worldwide as cold
beverages and as a hot drink-sour tea (Ismail et al., 2004). Roselle drinks are popular
under different names such as Karkade, ‘bissap’, ‘da bilenni’, ‘Jamaica’ and ‘Zobo’ in
different countries (McClintock and El-Tahir, 2004, Ismail et al., 2004 and Kolawole
and Maduenyi, 2004). According to Sayago et al. (2007), the intake of roselle flower
beverages in Mexican diet contribute to a consumption of 166 and 165 mg per serving
to the dietery fibre and polyphenols respectively. Olvera-García et al. (2008) reported
that hot aqueous extract of roselle (HAE) is used as refreshing beverages. Calyx is
utilized to colour and flavor the rum. Seeds are also used as an aphrodisiac coffee
substitute (Duke, 1983).
b. Fibre
Roselle produces a white and somewhat coarse bast fibre which is locally
used in Asia and West Africa for making twine cordage rope retting and sacks
(McClintock and El-Tahir, 2004). A strong silky fibre is extracted from the stems of
roselle and it is employed in Tamil Nadu by local people for cordage and manufacture

63
of gunnies (Maharashtra State Gazzetters, 1961). Some of the roselle fibre
characteristics are recorded in the Table 17.
Table 17. Proximate analysis and fibre dimensions of roselle fibres.
Sr. No.
Dimension/Component Quantity
1. Length (mm) 19-3.
2. Width (µm) 12-25
3. Cell wall thickness (µm) 3-7
4. Cold water solubility (%) 5.81
5. Hot water solubility (%) 8.24
6. Alcohol benzene solubility(1:2v v-1) (%) 3.88
7. 1% NaOH solubility(%) 25.80
8. Lignin(%) 19.20
9. Pentosans(%) 18.45
10. Holocellulose (%) 71.40
11. Hemicellulose(%) 22.80
12. α-cellulose(%) 48.60
13. β-cellulose(%) 9.56
14. γ-cellulose(%) 13.20
15. Ash (%) 1.40
16. Silica(%) 0.28
Source: McClintock and El-Tahir, 2004. Holocellulose: Hemicellulose+Cellulose
According to Crane (1949), average fibre yields run ca 1600 kg ha-1 with
yields in West Africa closer to 650 kg ha-1, 2100 kg ha-1 in Sri Lanka, 1500 in Java,
and experimental yields of 1200 to 3400 kg ha-1 in Malaya. If roselle is grown for
fibre much biomass remains as residue.

64
c. Paper and Pulp Industry
The bast fibres and stems of roselle are sometimes used for paper production
in USA and Asian countries (McClintock and El-Tahir, 2004). Khristova and Tissot
(1995) studied Soda anthraquinone pulping of roselle and Calotropis procera from
Sudan. Ahmed, et al. (1998 and 2000) studied delignification of roselle stem with
acetic acid, formic acid and propionic acid in organosolv pulping process. Saikia, and
Ali, (1999) reported Graft copolymerization of methylmethacrylate on to high alpha
cellulose pulp extracted from roselle.
d. Medicinal Uses
i. Role in Human Health
Essa and Subramanian (2007) reported that roselle is used in Ayurveda and
Traditional medicine in India, China and Thiland. The aqueous, methanolic and
ethanolic extracts of roselle plant are reported to play role in folk medicine (Dafallah
and al-Mustafa, 1996, Odigie et al., 2003, Adigun et al.,2006, Lans, 2006 and
Alarcon-Aguilar et al., 2007). Duke (1983) reported several medicinal uses of roselle.
Roselle is reported to have antiseptic, aphrodisiac, astringent, cholagogue, demulcent,
digestive, diuretic, emollient, purgative, refrigerant, resolvent, sedative, stomachic,
and tonic. Roselle is a folk remedy for abscesses, bilious conditions, cancer, cough,
debility, dyspepsia, dysuria, fever, hangover, heart ailments, hypertension, neurosis,
scurvy, and strangury properties. The calyx drink prepared in water, is used as a folk
remedy against cancer. Chen et al. (2003) reported that roselle is usually effective in
native medicines against hypertension, pyrexia and liver disorders. Kim et al. (2007)
stated that roselle a tropical beverage material and medicinal herb, is used commonly
as in folk medicines against hypertension pyrexia, inflammation, liver disorders and
obesity. Wright et al. (2007) in his review on ‘Herbal Medicines’, reported that roselle
plant has diuretic effects. Polysaccharides extracted from roselle flowers were helpful
to stimulate proliferation and differentiation of human Keratinocytes (Brunold et al.,
2004).
Different parts of roselle plant are used in traditional medicinal practices in
different countries. Philippines use the bitter root as an aperitive and tonic (Perry,
1980). In Brazil, stomachic, emollient and resolutive properties are attributed to the
bitter roots (Morton, 1987). According to McClintock and El-Tahir (2004), root

65
decoction of roselle can be used as luxative. McClintock and El-Tahir, (2004) have
listed several medicinal applications of leaves. Leaf extracts are often used
medicinally to treat colds, toothache, urinary tract infection and hangovers. Leaves are
applied as a poultice to treat sores and ulcers. In Senegal leaf juice is used to treat
conjunctivitis. Heated leaves are applied on the cracks in feet and boils and ulcers to
speed maturation. A lotion made from leaves is used on sores and wounds.
Medicinally, leaves are emollient and are much used in Guinea as a diuretic,
refrigerant, and sedative. Angolans use the mucilaginous leaves as an emollient and as
a soothing cough remedy. Central Africans poultice the leaves on abscesses (Watt and
Breyer-Brandwijk, 1962). According to Morton (1987), leaf infusions are used as
diuretic, cholectic, febrifuge and hypotensive and these also help to decrease viscosity
of blood. In East Africa the calyx infusion (Sudan tea) is taken to relieve from coughs.
Sharaf (1962) highlighted the hypotensive, antispasmodic, antihelmintic and
antibacterial activity of the calyx. Calyx infusions are used as a diuretic, cholerectic,
Febrifugal and hypotensive. It also helps to decrease viscosity of the blood and
stimulates intestinal peristalsis (Morton, 1987). Kirdpon et al. (1994) reported that
consumption of roselle juice in different concentrations and duration may helps to
treat and prevent the renal stone diseases. The seeds are diuretic and with tonic action.
The brownish yellow seed oil is claimed to heal sores on camels. Seed decoction is
used against dysuria, strangury and mild cases of dyspedpsia and debility (Morton,
1987). Ripe calyces are diuretic and antiscorbutic and fruits are antiscorbutic. Seeds
are reported to be diuretic and antiscorbic (Duke, 1983). In Burma, the seed are used
for debility and the leaves as emollient. Taiwanese regard the seed as diuretic,
luxative, and tonic (Perry, 1980). Leaf and Roselle calyx juice with salt pepper,
asafoeitida and Molasses, is taken as a remedy for biliousness.
ii. Pharmacological Studies
The antioxidant activity of roselle leaves and calyx has been demonstrated in
several experiments. It is attributed to presence of anthocyanins, Hibiscus
protocatechuic acid and polyphenolics (Duh and Yen, 1997, Tsai et al., 2002, Wong
et al., 2002 and Lee et al., 2009).
Tseng, et al. (1997) demonstrated protective effects of dried flower extracts of
roselle against oxidative stress in rat primary hepatocytes. Farombi and Fakoya (2005)

66
reported that dried flowers of roselle show free radical scavenging and antigenotoxic
activities of natural phenolic compounds. Prenesti et al. (2007) made antioxidant
power quantification of decoction and cold infusions of roselle flowers.
Protective role of roselle has been noticed by several workers against protein
degradation, lipid peroxidation and deformability loss of human erythrocytes due to
oxidative stress (Suboh et al., 2004). Amin and Hamza (2006) analyzed
hepatoprotective role of roselle against Azathioprine induced toxicity in rats.
Azathioprine inhibited the de novo and salvage pathways of purine synthesis. This led
to hepatotoxicity and oxidative damage. These problems could be relived with
administration of aqueous extracts of roselle. Further, they observed that the treatment
caused reduction in levels of MDA which could successfully release the inhibitory
effects of Azathioprine on catalase. Hirunpanich et al.(2005) reported that aqueous
extracts of dried calyces of roselle had antioxidant effects in CU mediated oxidation
of rat low density lipoproteins (LDL (p 0.05)). Hirunpanich et al. (2006) reported
hypocholesterolemic and antioxidant effects of aqueous extracts of the dried calyx of
roselle in hypercholesterolemic rats. Rasdhari et al. (2008) reported that incorporation
of roselle calyx extracts in yoghurt enhanced the total antioxidant property and
organoleptic qualities. Amin and Hamza, (2006) studied effects of roselle ethanol
extracts on cisplatin induced reproductive toxicity in (1g kg-1 26 days stage) rats. They
found that 10 mg kg-1body weight injection of roselle had protective effects and
treatment increased the activity of testicular antioxidant enzymes like GSH, SOD,
CAT etc. This also helped in restoring sperm mobility of cisplatin-treated rats.
According to them, protective effects of roselle might be mediated through their
potent antioxidant activities. Kao et al. (2009) reported that some polyphenol rich
extracts of roselle were beneficial in minimizing inflammation, including anti-
oxidative mechanism (enhanced catalase activity and glutathione) thereby improving
status under cancer and arthrosclerosis. It also reduced the liver problems such as lipid
peroxidation and liver lesions.
Kim et al. (2003) reported that H. sabdariffa extract inhibited the
accumulation of lipids and caused suppression of 373=L1 preadipocytes, adipogenic
transcription factors so as to block the adipogenesis in part by suppressing the
expression of C/EB p-alpha and PPAR gamma at protein levels. The rats were fed

67
during 4 weeks with either a basal diet, containing high cholesterol (1 %), cholic acid
(0.25 %), lard oil (10 %), or a supplemental diet with roselle extract at 5 %, 10 %, and
15 % levels (SD(5), SD(10), SD(15)). The significant decrease was noticed in LDL
level and triacylglycerol synthesis inhibition. Five percent roselle extract addition in
basal or supplementary diet showed the best results in the reduction of serum lipids
under study conditions. In Mexico, trials with Sprangue-Dawley rats with respect to
lipid profile showed that roselle extract decreased triacylglycerols and LDL levels in
all groups of treatment, however, total lipids were decreased at SD (10) and SD (15%).
There were significant alterations in triacylglycerols due to hibiscus acid recemization
from (+) –HCA to (-) – HCA in HSE treated Sprangue-Dawley rats (Carvajal-
Zarrabal et al., 2005). Liu et al. (2006) noticed protective effects of roselle extract on
CCl4 induced liver fibrosis in rats. Administration of the dried calyx extracts of roselle
at doses of 250, 500 and 1,000 mg kg-1 of the extracts significantly decreased
thiobarbituric acid reactive substances (TBARs) formation (p<0.05) (Hirunpanich et
al., 2006).The effect of roselle calyx extract on the fat absorption, excretion and body
weight in rats was studied by Carvajal-Zarrabal et al. (2009). The study indicated that
roselle extract can be considered as anti-obesity agent.
The flowers of roselle contain gossypetin, anthocyanin, and glucoside hibiscin,
which may cause diuretic and choleretic effects, decreasing the viscosity of the blood,
reducing blood pressure and stimulating intestinal peristalsis (Watt and Breyer-
Brandwijk, 1962). The effects of aqueous extracts of petals of roselle on kidney 1-Cl,
renovascular hypertension were investigated by Odigie et al. (2003). 250mg.kg-1.day-1
dose in drinking water was able to reduce BP. Haematological investigations showed
that dose dependent oral administration was beneficial to enhance Haemoglobin,
haematocrit but not haemoglobin.
Onyenekwe et al. (1999) observed significant decrease in serum creatinine,
cholesterol and glucose in roselle calyces infusion treated spontaneously hypertensive
rats. According to Faraji and Tarkhani (1999), roselle sour tea is reported to have
properties of lowering of high blood pressure and suggested more extensive studies on
this subject. Odigie, et al. (2003) reported that chronic administration of aqueous
extract of roselle attenuated hypertension and reversed cardiac hypertrophy in 2K 1C
hypertensive rats. Hibiscus sabdariffa Extract (HSE) significantly reduced foam cell

68
formation and inhibited smooth muscle cell migration and calcification in the blood
vessels of rabbits. The results showed that HSE inhibited serum lipids and showed an
anti atherosclerosis activity (Chen et al., 2003). Chen, et al. (2003) reported that
roselle extract inhibited the development of atherosclerosis in cholesterol fed rabbits.
Herrera-Arellano,et al.(2004) studied effectiveness and tolerability of a standardized
extract from roselle in patients with mild to moderate hypertension in a controlled and
randomized clinical trial. They reported a cure of mild and moderate hypertension
with roselle extracts infusion. It was proved to be a significant antihypertensive drug
(at 9.6 mg anthocyanin per day (from 10g of dried calyces on 0.5% water) daily
before breakfast. Lans (2006) in his survey of some medicinal plants noticed that
roselle is used against hypertension. Mojiminiyi et al. (2007) observed
antihypertensive, hypotensive and negative chromotropic effects in rats after
application of aqueous calyces extract of roselle on two types of experimental
hypertension, salt induced and L- NAME (NC0mega)- L- arginine methyl ester)
induced in permosensive controls. Iyare and Adegoke, (2008) showed that maternal
consumption of aqueous extracts of roselle during lactation decreased postnatal
weight gain and delayed onset of puberty in the female offspring. Mozaffari-Khosravi
et al. (2009) reported antihypertensive effectiveness of sour teat (ST-H. sabdariffa)
infusion in diabetic patients after consumption of ST infusion. Lee et al. (2009)
recommended polyphenolics rich extracts of roselle for neutropathy in diabetes. Ojeda
et al. (2009) showed inhibitory action of roselle extract on activities of ACE
(Angiotensin Converting Enzyme). This observation supports the data related to folk
medicine use of roselle calyces as antihypertensive agent.
A low dose of roselle juice was found beneficial than a high dose in renal
stone diseases (Kirdpon et al., 1994). Chen et al. (2004) reported inhibitory effects of
roselle extract on low density lipoprotein oxidation and anti hyperlipidemia in
fructose fed and cholesterol fed rats. In human beings also better results were obtained
against urinary problems. Herbal infusion of roselle significantly reduced the
creatinine, uric acid, citrate, tartarate, Ca, Na, K and phosphate in the urine. But it did
not affect the reduction of oxalates (Ali et al., 2005). Orisakwe, et al. (2004) reported
testicular effects of sub chronic administration of roselle calyx aqueous extract in rats.

69
Lin et al. (2002) reported anti-hepatoma activity of fifteen natural medicines
(including roselle) from Canada on liver cancer cell lines and indicated that the crude
drugs are highly effective against hepatitis B virus having cytotoxic properties.
Chewonarin, et al.(1999) studied the effects of roselle on the mutagenecity of
various known mutagens in Salmonella typhimurium and on formation of aberrant
crypt foci induced by the colon carcinogens azoxymethane and 2 amino 1 methyl 6
phenylimidazo (4, 5 b) pyridine in F 344 rats. They noticed that at the initiation stage,
the number of AOM-induced ACF in the colon was significantly decreased by roselle
(17-25 %) compared with that in rats treated with AOM alone. The amount of O6-
methylguanine in the colonic mucosa was found to be decreased in the roselle-treated
rats. The number of PhIP-induced ACF was also significantly decreased by roselle
treatment (22 %) at a concentration of 1.0 g kg-1 body weight in the initiation stage.
However, in the post-initiation stage of AOM-induced ACF formation, roselle
increased the number of ACF, especially the number of foci which had more than
three crypts/focus. These results indicate that roselle has antimutagenic activity
against MAM acetate and heterocyclic amines and that it decreases the number of
AOM- and PhIP-induced ACF in the initiation stage, although it rather increased the
number of ACF in the post-initiation stage.
Fakeye et al.(2008) have stated that there was significant decrease in onset of
sleep and increasing in sleeping time after a dose of system at A300, AE50 and
AE300 mg kg-1 doses groups (P<0.001; P<0.01 and P<0.05 respectively). However,
ethanol extract had slow impact on sleeping time and some doses (EAC at 50 mg kg-1
= with P<0.05) is beneficial in reduction of sleeping time. This observation reveals
anxiotypic and sedative effects of roselle extract.
Asagba et al.(2007) reported protective role of aqueous extract of roselle petal
against chronic exposure to Cd after study of some selected biochemical parameters in
12 week old male wistar albino rats. It also protected against Cd induced liver
prostate, and testes lipid peroxidation as evidenced by significantly reduced MDA
values in these organs.
Ali et al. (1991) investigated the antispasmodic potential of roselle calyces.
The succulent calyx, boiled in water, is used as a drink in bilious attacks (Watt and
Breyer-Brandwijk, 1962).

70
Chua et al. (1987) studied antimicrobial properties of roselle. They reported
that the antimicrobial property of roselle calyx extracts (aqueous and ethanol) was not
affected by heat treatments. However, in combination with protocatechuic acid,
ethanol extract of roselle calyx exhibited dose dependent inhibitory activity. The
ethanol extracts of roselle calyx exhibited greater antibacterial effects than the
aqueous extract. They suggested that in ethanol extract of roselle calyx,
protocatechuic acid might became potent agents as food additives to prevent
contamination from the bacteria. Rukayadi et al. (2008) assessed thi medicinal plant
(H. sabdariffa) for their anticandidal activity against six Candida species viz. C.
albicans, C. glabrata, C. guilliermondii, C. Kruisei, C. parapsilosis and C.
tropicallis. They found that methanolic extracts of roselle fruits had low anticandidal
activity (range of 100-500 mg ml-1).
Rukmini et al. (1982) made toxicological evaluation of roselle oil. The oil has
cyclopropenoid fatty acids (2.9 %) and epoxy fatty acids (2.6 %) along with normal
fatty acids in vegetable oil. Rats fed with this oil showed slower growth and inferior
reproductive performance with some alterations in liver metabolism. From the acids
identified from this oil, sterculic acid (C19) and malvalic acids (C18) are toxic one. It
is evident from this report that the roselle seed oil in raw state is unsafe for human
consumption.
e. Other Uses
The seeds are used as feed for chickens. Roselle cake is used as cattle feed
when available in large quantity (Morton, 1987). The leaves are a source of mucilage
and are used in pharmacy and cosmetics (McClintock and El-Tahir, 2004). According
to Wiam et al.(2006), roselle can be used as a histological stain. Sato et al. (1991)
studied structure and contents of main coloring constituents in the calyces of roselle
and commercial roselle color. Recently, the ornamental use of roselle as a garden
plant, as cut flowers is under consideration (McClintock and El-Tahir, 2004). The red
stalks with ripe red flowers are used in flower arrangement in Europe.

71
C. About Hibiscus tiliaceus Linn.
1. Introduction
Although Hibiscus is a quite large genus of family Malvaceae, only one
Hibiscus species shows distinct halophytic traits and this species is a native of Pacific
Islands and it is cultivated in the tropical and subtropical belt. It is an important
component of mangrove ecosystem in India. The plant is having a second value in the
ancient cultures of Hawaii and Tabiti Islands and it was so highly valued that
permission to cut it was to be obtained from the village chief (ntbg.org/plants/details
php ? plastid 6255).
Hibiscus tiliaceus L. belongs to section Azanzae DC of family Malvaceae
(Fryxell, 1988). On the basis of leaf shape and degree of marginal lobbing 5
subspecies were recorded (Borassaum Waalkes, 1966), but according to Elevitch and
Thomson (2006), there are 7 subspecies namely H. tiliaceus subsp. tiliaceus
(widespread and may have variegated leaves, double flowers, white petals with
maroon spots at the base, and pure yellow flowers are known forms) (Staples and
Herbst, 2005), H. tiliaceus subsp. elatus (Hochr.) Borss. Waalk. Lime tree leaved
Hibiscus, H. tiliaceus subsp. genuinus (Hochr.) Borss. Waalk. H. tiliaceus subsp.
hastatus (L.f.) Borss. Waalk. (from Pacific islands with three lobed leaves) H.
tiliaceus subsp. similis (Blume) Borss. Waalk. (native to Southeast Asia. Rerely with
mature capsules and may have evolved as a hybrid with H. macrophyllus), Hibiscus
tiliaceus subsp. celebicus (Koord.) Borss. Waalk. (only from Sulawesi, Indonesia)
and Hibiscus tiliaceus subsp. crestaensis Borss. Waalk. (only from the Island of
Luzon, Philippines)
There are different synonyms for Hibiscus tiliaceus. These are as follows H.
tiliaceus var. heterophyllus Nakai, H. tiliaceus var. tortuosus (Roxb.) Masters, H.
tortuosus Roxb., H. boninensis Nakai, H. celebicus Koord., H. hastatus L. f., H.
similis Blume, Paritium elatum (Sw.) G. Don, P. simile (Blume) G. Don, P. tiliaceum
(L.) A. Jussieu, P. tiliaceum var. heterophyllum (Nakai) Nakai, Pariti boninense
(Nakai) Nakai, P. grande Britt. ex Small, P. tiliaceum (L.) A. Juss. ex Britt. and
Millsp., P. hastatum (L.f.) O. Deg. and Greenwell,Talipariti tiliaceum (L.) Fryxell T.
celebicum (Koord.) Fryxell, T. crestaense (Borss. Waalk.) Fryxell, T. elatum (Sw.)
Fryxell, T. hastatum (L. f.) Fryxell, T. simile (Blume) Fryxell, T. tiliaceum var.

72
pernambucense(Arruda)Fryxell(httl://zipcodezoo.com/plants/H/Hibiscus_tiliaceus/def
ault.asp;http://tropicos.org/Name/50211911,http://tropicos.org/NamePrint.aspx?Name
id =502 11759 and http://www.tropicos.org/Name/50211761).
Some of the vernacular names for H. tiliaceus are mentioned in Table 18.
Table 18. Vernacular names of Hibiscus tiliaceus L.
Language Vernacular Names
English
Coast Cotton Tree, Yellow Mallow Tree, Beach Hibiscus, Sea Hibiscus, Malabago, Majagua,
Mahoe, linden hibiscus Coast cottonwood, Cuban bast, Hau, Bladder Ketmia, Cotton Tree,
Cottontree, Hawaiian Tree Hibiscus, Lagoon Hibiscus, Linden Hibiscus, Mahoe, Majagua,
Mountain Mahoe, Sea Hibiscus, Tree Hibiscus
Sanskrit Bala
Hindi and Bengali Bola, Chewla,
Marathi Belpata
Telugu Etagoga
Tamil Nirparathi,
Malyalam Attaparathi
Oriya Baniah
Hawaii “O-hamabo” “Hau Uacimada Praia”, hau
Madagascar Aviavy
Source : Watt, (1890), Sastri, (1959), Cooke, (1967 b) and Tanaka (1976).
Besides being an important component of mangrove ecosystem in the tropical
world, H. tiliaceus shows great environmental adaptability. It usually occurs at low
elevation and may be up to 800 m (2600 ft.) (Konczak, 1996 and Saquet, 1996) with
annual rainfall 900-2500 mm (35-100 in.) and at mean annual temperature 12-32 0C
(54-90 0F) maximum temperature of 24-41 0C (75-106 0F) and minimum temperature
of about 5-24 0C (41-75 0F), minimum tolerated limit 0-14 0C (32-570F) of
temperature and 0-6 months of dry season i.e. 40 mm (1.6 inch). In coastal

73
ecosystems where substrate salinity is relatively high, some times, it occurs near
ocean, streams and in moist sloping areas, it inhabits fresh water to saltwater
(http://pdf.usaid.gov/pdf_docs/PNADN809.pdf). Best tree growth occurs in full sun. It
can grow well in light to heavy textured soils, sandy loams, loams, sandy clay loams,
sandy clays and clay loams (Shazra et al., 2008). It grows in soils with free or
impeded drainage. It tolerates acid to alkaline soils (pH 5.0 to 8.5). It is also occurs at
high pH sites It is tolerant of brackish conditions and shallow water flooding for many
months. Uniform rainfall patterns and summer are favourable for its growth; the tree
can be grown in waterlogged soils, marsh and limestone’s (Allen, 2002).
2. Distribution
Tropical and subtropical regions of Pacific islands are rich in this species as it
has its origin on the same islands (Whistler, 1980, Chin and Enoch, 1988 and
Konczak, 1996). The plants of H. tiliaceus L. are found along the Eastern and
Western coast of India near backwaters and banks (Cooke, 1967 b and Pattanaik et
al., 2008). It is the most common undergrowth species of Sundarban Mangrove forest
and is dominated all over forest (Rashid et al., 2008). In Andaman Islands, it is found
to form dense thickets near the shore. The plants are also noticed in Konkan:
Ratnagiri banks of Terrecol river Kanara: along the sea-coast and banks of tidal rivers
(Cooke, 1967 b). Hibiscus tiliaceus a typical mangrove associate, appears to have
great phenotypic plasticity. This is demonstrated in Hawai the most isolated
archipelago on earth. In this island group H. tiliaceus has been able to expand its
habitat range into wet montane forest zones upto 1200m in elevations (Wagner et al.,
1990). Ridley, (1930) noticed that the floating ability of H. tiliaceus seeds due to air
spaces between the folds of cotyledons. He noticed that after floating for 40 days in
sea water, the seeds fail to germinate in sea water. The dispersal ecology of 17
maritime species was studied by Nakanishi (1988). He estimated that Hibiscus seeds
has got 14. 1 mg dry weight, 0.5 cm3 volume and 0.28 g cm2 density due to air spaces
between folded cotyledons. He noticed that seeds started to sink after 1-5 weeks but
number of seeds (> 50 %) remained floating for 2-3 months. He further observed
increase in germination percentage due to floating in seawater. Nakanishi (1988)
indicated that drift disseminules on drift line seedlings on the high tide line in herb

74
zone. Absence of young plants in the herb zone were the characteristic feature of H.
tiliaceus in the Ryukyu Island of Japan.
3. Botany
Hibiscus tiliaceus is a evergreen shrub or small tree or Tree; about 10-20
meters tall. Leaves are 4-5 inches in length, cordate roundish ovate, abruptly
acuminate, finely reticulate veined, entire or crenullate; leathery, with smooth edges,
while the underside is velvety and with an indumentum white, with very short, stellate
hairs, occasionally the leaf edges are scalloped. Leaves usually with 9-11 main veins
and always 1-3 glands on the abaxial surface near the petiole insertion (Tomlinson,
1986); petioles 1½ - 2 inches long; with a pair of large, grey green, leafy stipules,
oblong in shape, subulate. A dense cover of white, very short, stellate (star shaped)
hairs on thestalks and lower surface of the leaves makes these parts appear grey.
Flowers are produced throughout the year singly in the axil of the leaves, on
the base long peduncle enclosed by the stipule pair, stipules fall later and leave a
conspicuous annular nodal scar (Tomlinson, 1986). Flowers are 5-7 cm in diameter,
Calyx 3/8 inch long , divided nearly to the base ; lobes narrow -lanceolate with a
prominent mid nerve, tomentose, calyx conspicuously 5 toothed, persistent in fruit
(Cooke, 1967 b and http://pdf.usaid.gov/pdf_docs/PNADN809.pdf). Glands present on
the back of the epicalyx lobes but appear to be functionless in the act of pollination.
Corolla 2/3 inch across, pubescent out side, yellow in the morning, turning red in the
afternoon with dark crimson centre. Flowers usually with a continuous maroon eye;
receptacle without yellow latex. The surfaces of the petals and stamen tube are
covered with glandular hairs. Staminal tube antheriferous all the way down styles
connate 3/16th inch below the stigmas. Pollen fertility is 100%.
Floral formula:
Fruit is a capsule½ inch in diameter, hairy, globose with a short beak, ovoid in
shape, light brown in colour and about 2 cm long. Spuriously l0 celled, 5 valved
hidden in the persistent calyx, tomentose. Capsule regularly loculicidally dehiscent
and it splits into five segments upon drying gaping widely. (Borassaum Waalkes,
1966; Tomlinson, 1986; Selvam, 2007 and https://www.epa.qld.gov.au/register/p00

75
273aa.pdf). The seeds are exposed when the capsule splits; Seeds are 4mm long,
smooth kidney shaped to suberiform, brown to brown-black, sulcate (Selvam, 2007).
It is noticed by Bell (2006) that leaf width and length, petal width and length,
burgundy centre colour and anther counts are significantly different in plants growing
in coastal region and mountain region.
4. Anatomy
The heartwood of H. tiliaceus is red and fades on exposure to light
(Anonymous, 1956). Wood anatomy of the tribe Hibiscae with respect to leaf and
sporoderm anatomy and ontogeny of secretary structures were studied by Rocha et al.,
(2002). Silva, (1966) studied pollen structure in H. tiliaceus and H. Pernambucensis.
Anatomical characteristics of the seeds are illustrated with emphasis on seed coat
structure and development (Soave, 1990).
5. Cytogenetical Studies
Hibiscus tiliaceus has chromosome number 2n=96 (Krishnappa and
Munirajappa, 1980 and Cheng and Tsai, 1999). In China nine natural populations
have been studied to investigate genetic variation with the help of AFLP within and
among the populations of H. tiliaceus. Polymorphism was absent but long distance
dispersal of floating of seeds and the local environment play a major role in genetic
diversity and genetic structure of this species. (Tang et al., 2003). Overall study
indicates that the species is characterized by a relatively high genetic diversity within
population.
According to Tang et al. (2003), high genetic diversity indicates adaptability
of the H. tiliaceus to a wide range of environmental changes. This reflects the current
adaptive capacity of H. tiliaceus to various habitats (Hamrick et al., 1979 and Ledig,
1988). Wu et al. (2007) studied PCR mediated recombination of the amplification
products of the H. tiliaceus L. glyceraldehyde-3-phosphate dehydrogenase gene. Yang
et al. (2008) performed isolation and high quality total RNA extraction from H.
tiliaceus L a mangrove associate and its relatives. According to Takayama et al.
(2005), H. glaber a woody species is derived from H. tiliaceus. It is mostly found on
Bonin island of Northern West Pacific Ocean. According to their cpDNA analysis,
haplotypes are divided into 6 sections (Bonin, Kazan and Ryukyu islands while,

76
Okinawa, Kagoshima, Mie and Aichi prefectures are in the Japanese archipelago,
southeast Asia and Pacific Island) including 37 types of same localities, which are
further divided into 13 haplotypes Type B , Type J, Type K, Type L, Type M, Type
N, Type O, Type P, Type Q, Type R, Type S, Type T and Type U4’ 5 (Takayama et
al., 2005). The Western North Pacific Oceanic Islands showing occurrence of H.
tiliaceus are mentioned below Bonin Islands; Chichijima Islands, Hatsuneura,
Chichijima I., Kominatokaigan, Chichijima I., Butakaigan, Chichijima I., Nakakaigan,
Chichijima I., Higashijima I., Bonin Islands;, Hahajima Islands, Anejima I., Bonin
Islands; Mukojima Islands- Mukojima I., Mukojima I.,Nakôdojima I.,Kazan Islands-
Kita-Ioujima I., Okinawa Pref.; Ryukyu Islands, Tamashiro Vill., Okinawahonto I.
,Ada, Okinawahonto I., Ikeijima I., Akajima I., Okinawa Pref.; Miyako Islands,
Shigira beach, Miyakojima I., Sawadanohama, Irabujima I., Shimojijima I., Okinawa
Pref.; Yaeyama Islands, Ishigakijima I., Ohta, Ishigakijima I., Taketomijima I.,
Haimida, Iriomotejima I., Haimida, Iriomotejima I., Haterumajima I., Kagoshima
Pref., Kasari Town, Amamiohshima I., Southeast Asia-Labrador Park, Singapore-
Noordin, Ubin, Singapore, Pelabuhanratu, Java, Hua Thanon, Samui, Phuket, near
Laem Sai, Noordin, Ubin, Singapore, Dodanduwa, Sri Lanka ,Taiwan, Pacific Islands,
Saipan I., Saipan I., Hawaii I., Hawaiian Is., Kauai I., Hawaiian Is., Faleolupo and
Savaii I. Takayama et al. (2005) studied phytogeography and genetic structure of H.
tiliaceus L. from sea drifted seeds. According to their cpDNA analysis H. tiliaceus is
the species which is responsible for the recurrent speciation of all the allied species of
H. tiliaceus (Takayama et al., 2006).
6. Physiological Studies
Ng and Asri (1979) recorded that there is physical dormancy in seeds of H.
tiliaceus which was due to impermeable seed coat. Scarification of seed coat with
sand paper and by nicking with a knife is found suitable for ensuring timely
germination (Allen, 2002). Annual growth rate in H. tiliaceus is estimated at 0.75 to
1.5 m.year-1 in height, which indicates that H. tiliaceus is a fast growing tree (Elevitch
and Thomson, 2006). Criley (1997) reported that vegetative growth of H. tiliaceus
was retarded by foliar sprays and drench applications of chloromequat, uniconazole,
paclobutrazol and fluprimidol. Among the four compounds chloromequat was most

77
effective. The major physiological studies performed in H. tiliaceus are pertinent to
salinity tolerance. This pantropical mangrove species usually occurs in coastal
ecosystems but it also inhabits upland habitats in Hawaii. Santiago et al.(2000) raised
the plants from cuttings from three populations on the Islands of Oahu, Hawaii, in
glasshouse under two levels of substrate salinity (0 and 335 mOsm kg-1) and three
light treatments (0 %, 50 % and 90 % shade). Light had stronger effect on leaf area
ratio (LAR) and leaf mass per area (LMA), whereas salinity had stronger effect on
leaf water content, internode length, and plant biomass. Salinity reduced total new
biomass by 58, 50, and 34 % in full sun, 50 % and 90 % in shade respectively, but this
response did not differ between populations. It was further noticed that salinity
reduced net CO2 assimilation in upland population but had no effect or stimulated
photosynthesis in coastal populations. Increasing salinity decreased stomatal
conductance in all populations and therefore increased water-use efficiency. The
degree to which photosynthesis was inhibited by salinity was inversely proportional to
the salinity of source population, indicating a loss of salinity tolerance in upland
plants. Salinity reduced photosynthesis but not growth, of upland plants because
increased allocation to photosynthetic tissue increased LAR to compensate for
inhibition of photosynthesis by salinity. Naidoo et al. (2002) studied changes in gas
exchange and chlorophyll fluorescence characteristics of H. tiliaceus L in response to
salinity in the natural environment (at low salinity site Beachwood <12 permill and
high salinity site Durban Bay 35 permill.). These involved measurement of CO2
exchange, chlorophyll fluorescence, and ion and water relation at two sites. At
Beachwood, CO2 exchange was highest (9.12 µmol m-2s-1) and at the Durban Bay
site, CO2 exchange decreased than that at Beachwood by 59 %. At saturating PFD,
PSII quantum yield and ETR (Electron Transport Rate) were lower at high salinity
site. Photochemical and non-photochemical quenching was higher at the high salinity
site. There were no differences in ion concentration between sites except for K+,
which was significantly higher at Durban Bay. Eventhough H. tiliaceus is a mangrove
associate at salinities ≥ 40 %, plants showed complete defoliation followed by 100 %
mortality within 1 week (Aronson, 1989, Yossef, 1995 and Santiago et al., 2000).
Total Methylated Onium Compounds (TMOC) are important osmolytes since
they are present in appreciable amount (101.6 mol m-3 plant water) in this halophytes

78
species (Popp et al., 1984). In young leaves accumulation of mannitol was noticed
while in old leaves mannitol and sucrose were predominant. The species showed
K/Na ratio greater than unity. Popp (1984) noticed that old leaves accumulated more
sodium, calcium, citrate and malate than young leaves while opposite trend was
noticed in case of chloride, sulphate, potassium and magnesium. Soto, (1992) studied
nutrient retranslocation during leaf senescence. His study revealed that there was
26.6 % decrease in potassium content and 4.78 % increase in magnesium level in
senescent leaves of this species. A very negligible extent of salt excretion from H.
tiliaceus leaves in comparision tosalt secreting mangroves such as Avicennia and
Aegiceras was evident in the experiments of Scholander et al. (1962).
Besides salinity, mangroves and mangrove associates growing in estuarine
areas are frequently exposed to waterlogging stress. Under flooding conditions, in the
soil, the roots of sensitive species generally change their respiratory mode from
aerobic to anaerobic and show accumulation of harmful products like ethanol and
lactic acid. During ten days anoxic treatment, no ethanol accumulation in seedlings of
H. tiliaceus was noticed indicating a good waterlogging tolerance potential (Yossef,
1995).
Shade grown and sun grown H. tiliaceus plants were analysed for composition
of carotenoids with the help of HPLC technique. Shade grown H. tiliaceus contains
23-76 % of total carotenoids whereas, sun grown seedlings showed considerable
decline (Thayer and Bjorkman, 1990). Youssef and Saenger, (1998) noticed
accumulation of sulfur in plants of H. tiliaceus at low irradiance. Desai et al. (2010)
studied antioxidant potential of H. tiliaceus L. leaves from west coast of Maharashtra
locality Ratnagiri. It is revealed that young leaves of H. tiliaceus L. are a better
candidate among ten coastal plant species studied for their DPPH radical scavenging
activity, reducing power, total polyphenol and flavonoid contents.
7. Phytochemical constituents
Analysis of H. tiliaceus leaves by different workers revealed following
constituents.
Protein : 11.8 %
Fat : 4.8 %

79
Starchy matter : 52.1%
Crude fibres : 21.5 %
Ash : 9.9 %
Values are in percent dry weight.
(Akinwusi and Illoh, 1995; Chun, 1995, Field, 1995, Allen, 2002 and Bell, 2006)
The heartwood of H. tiliaceus is red and fades on exposure to light. Study of
naturally occurring compounds revealed presence of hibiscoquinones in the
heartwood (Anonymous, 1956). It contains hibiscones A–D and hibiscoquinones A–
D; and wood sample which is devoid of red pigments showed presence of lapachol
(Ali et al., 1980). The roots of this species contains (from Brazil) gossypol, and
mansonones D and F (Akinwusi and Illoh, 1995).
A new coumarin, hibiscusin, and a new amide, hibiscusamide, together with
eleven known compounds including vanillic acid, P-hydroxybenzoic acid, syringic
acid, P-hydroxybenzaldehyde, scopoletin, N-trans-feruloyltyramine, N-cis-
feruloyltyramine, a mixture of beta-sitosterol and stigmasterol, a mixture of beta-
sitostenone and stigmasta-4,22-dien-3-one were isolated from the stem wood of H.
tiliaceus by Chen et al. (2006). Along with this a new friedelane-type triterpene
named 27-oic-3-oxo-28-friedelanoic acid together with eight known triterpenoids
involving five friedelane-type derivatives were isolated (Li et al., 2006).
Hydroxybenzoic acid Vanillic acid Syringic acid
β-sitosterol Stigmasterol Stigmasterin

80
Fig. 1. Secooleanane and its derivatives from H. tiliaceus L.
The 3, 4-secooleanane derivatives 716–719 from the mangrove plant H.
tiliaceus (335) have been identified (Chen et al., 2006). The secoolea-nanes 718 and
719 are shown in the reference with the 18a-configuration but there is no evidence for
this stereochemistry (Fig. 1). Other new lanostanes include the nigrumane derivatives
42–44 from H. tiliaceus are in Fig. 2.
Fig. 2. New lanostanes and nigrumane derivatives from H. tiliaceus L.
Rosa et al. (2006) reported that methanolic flower extract of H. tiliaceus
contain Vitamin E and some stigma sterol derivatives. Melecchi et al. (2002)
optimized the sonication extraction method of H. tiliaceus L. flowers and found some
saturated hydrocarbons, fatty acids, fatty acid methyl esters and vitamin C. these
phytochemicals were identified on GC/MS analyzer. Crude protein analysis with
electrophoresis was carried out from seeds of some Hibiscus species including this
particular species (Akinwusi and Illoh, 1995).
8. Cultivation
Hibiscus tiliaceus is a multipurpose tree with ecological and ornamental value.
Hence, attempts are made to standardize the cultivation of this species. The
propagation of plant can be achieved by both seed germination and vegetative
propagation. But it is a common experience that this is rather a difficult operation
with about 50 % success (Elevitch and Thomson, 2006). The scarification of seeds

81
with sand paper and nicking of the seed coat with knife or nail clipper is
recommended to promote seed germination which occurs within 2-4 weeks. It is
indicated by Elevitch and Thomson (2006) warm, moist and light conditions are
favourable for optimum germination in trays filled with peat and perlite or vermiculite
(Nakasone and Rauch, 1973) recommended transplantation of 5-6 months old
seedlings of about 25 cm height are transferred to field or out-planting was done.
Propagation with branch cutting is the most common method of propagation in
case of this species. For development of fences 2-3 m cuttings of straight branches are
taken they buried during rainy season or wet weather, these are buried ½ to 2/3 of
their length deep into the moist soil (Elevitch and Thomson, 2006). For direct sowing
as well as vegetative propagation rainy season is most favourable. Once established
little maintenance is required but, according to Nagata (1992) proper pruning and
aftercare is essential to check irregular spread of the tree. The plant is relatively
resistant to pest and diseases (Elevitch and Thomson, 2006).
9. Uses
a. Human and Animal Nutrition
In Java young leaves are fermented into sauce and consumed (Tanaka, 1976).
Young leaves are eaten in Java, where they are fermented into sauce. Australian
aborigines eat the roots (Tanaka, 1976). Ethnobotanical studies of the tribe of
Andaman and Nicobar Islands India. Seeds are eaten after roasting fruits (Bhargava,
1983). Leaves bark and roots are preferred as famine food (Curl and Truckove, 1969;
Chopra, et al., 1973; Dey, 1973; Agarwal and Ghosh, 1985; Chin, 1992; Chun, 1995;
Trivedi, 1997, Elevitch and Thomson, 2006 and https://www.epa.qld.gov.au/regis
ter/p00273aa.pdf). H. tiliaceus is included in the group of major honey plants of
Sundarbans (Prain, 1903). In Malaysia, leaves are utilized as cattle feed (Akinwusi,
and Illoh, 1995) (Plate 2).
b. Medicinal Uses
In the Ethnology Collection at Bishop Museum (Hawaii) there is exhibit of
bowl made from wood of H. tiliaceus (Brown, 1935). Water kept for 3 days in a
container made of belpatter wood (H. tiliaceus L.) is administered orally for diabetic
in Kurukhetra district Haryana India (Lal and Yadav, 1983). In Fiji the leaves are

82
wrapped around limb fractures and the stem is used as part remedy for treating ulcers
(Smith, 1981). It is indicated that leaves may expel salt and toxins, to protect the inner
tissue from extreme isolation (Karimi and Ungar, 1989). In Bangaladesh decoction of
leaves is used as hair restores, expectorants and for treatment of obstinate causes of
urine. (WWW SOS-arsenic net/English/homegarden/index html).
Lotion of infusion of leaves is applied for ulcers and wounds. The leaves are
also considered as laxative and resolvent (Pattanaik et al., 2008). The leaf buds are
chewed/swallowed for dry throat. The bark is emetic. The mucilage, obtained by
macerating fresh bark in water, is given in dysentery. The inner bark (with sap) is
soaked and drunk for relief from labor pains and rubbed on stomach. Sap used as an
internal lubricant as a mild laxative and to facilitate the passage of a foetus through
the birth canal (Bourdy and Walter, 1992). Hawaian used the slimy sap from the inner
bark as a laxative. The root is febrifuge, aperitive, emollient, sudorific diuretic and
laxative; it is used in the preparation of embrocation for rheumatism and lumbago
(Han, 1998). Flowers are boiled in milk and used as a remedy for earache. The
flowers are thought to aid in indigestion.
i. Pharmacological Studies
Rosa et al. (2006) studied pretreatment effect of H. tiliaceus L. methanolic
extracts (HME) on V79 cells against cytotoxicity and genetotoxicity induced by
hydrogen peroxide and tert-butyl-hydroperoxide. They proved that HME is with
antioxidant activity and it had antimutagenic and antotoxic effect against oxidative
DNA damage. Methanolic extracts of H. tiliaceus L. showed no toxicity against
fibroblast but selective cytotoxicity against breast cancer cell (Ic (50), 1.1-1.6 mg ml-
1) (Shaikh et al., 2011). Chen et al. (2006) reported that a novel amide hibiscusamide
from stem wood of H. tiliaceus exhibited cytotoxicity against P-388 and/or HT-29
cell lines in vitro. Rosa et al. (2006) reported that treatment of methanolic extract of
flowers protected several Saccharomyces cerevisae strains defective in antioxidant
defenses against H2O2 and t-BooH cytotoxicities showing a clear antioxidant activity
of flower extract. These workers suggested that this effect may be due to
nonenzymatic antioxidants such as vitamins and phytosterols. It was further noticed
that the extract was not mutagenic and it showed a significant antimutagenic actions
against oxidative mutagens in S. cerevisae. Mubassara et al. (2011) recently noticed
that alcohol extract of H. tiliaceus had polyphenols and antioxidant activity and it

83
inhibited release of histamine and leukotriene B4 (LTB4) induced by the calcium
ionophore A23187 from rat peritoneal exudates cells. According to these workers this
may beee useful in development of antiallergic supplement and drugs. The
experiments of Masuda et al. (2005) revealed that the extract of H. tiliaceus had
significant tyrosinase inhibitory activity.
c. Other Uses
Wood is buoyant and light in weight, but stronger land better suited for larger
canoes (Elevitch and Tomson, 2006). Its wood is used as fire wood, wood for boat
making, Cabinet and for fancy work (Pattanaik et al., 2008). Stems are utilized for
ground poles and fuel wood in Northern, Central and Southern regions of Vanuatu
Country. The bark yields a strong fibre used in Bengal for ropes and cordage. In Ceylon
it is used for mats (Maharashtra State Gazetteers, 1961).
Hibiscus tiliaceus plays a role in soil erosion control and coastal land and sand
dune stabilization. The ornamental value of the H. tiliaceus trees in modern landscapes
is recognized. The species is often planted as supporter and shade tree for piper bettle
in south East Asia. (Cooke, 1967 b). Evaluation of regenerated deforested sites of
coastal windbreaks revealed that amongst 12 tree species H. tiliaceus L. is the best
with height growth of 4.5 m and diameter, 7.76 cm (Kan and Hu, 1987). In secondary
forest it is found to be an important colonizing species (Webb et al., 1999). It is
evident from the foregoing account that commercial potential of H. tiliaceus is not
completely understood and fully exploited especially in country like India.

![Defatted Kenaf (Hibiscus cannabinus L.) Seed Meal and Its ...downloads.hindawi.com/journals/omcl/2018/6742571.pdf · [18, 23, 24]. The recoveries of phenolics and saponins in PSRE](https://static.fdocuments.net/doc/165x107/5fdc4a51e086324e234137c5/defatted-kenaf-hibiscus-cannabinus-l-seed-meal-and-its-18-23-24-the.jpg)
















