
3 Infectious Diseases of Coldwater Fish in Marine and ... · Clinical signs of IPN in post-smolt...
45
3 Infectious Diseases of Coldwater Fish in Marine and Brackish Water Michael L. Kent 1 ,* and Trygve T. Poppe 2 1 Department of Fisheries and Oceans, Biological Sciences Branch, Pacific Biological Station, Nanaimo, British Columbia V9R 5K6, Canada; 2 Department of Morphology, Genetics and Aquatic Biology, The Norwegian School of Veterinary Science, PO Box 8196 Dep., N-0033 Oslo, Norway Introduction Salmonids are the primary fishes reared in cold seawater netpens. This component of the industry produces approximately 500,000 t year −1 on a worldwide basis. The principle species reared in netpens are Atlantic salmon (Salmo salar), coho salmon (Oncorhynchus kisutch), chinook salmon (Oncorhynchus tshawytscha) and rainbow trout (Oncorhynchus mykiss). Additional species include minor produc- tion of Arctic char (Salvelinus alpinus), Atlantic cod (Gadus morhua), haddock (Melanogrammus aeglefinus), Atlantic halibut (Hippoglossus hippoglossus) and Atlantic wolffish (Anarhichas lupus). The purpose of this chapter is to review the most important infectious diseases affecting fish reared in cold seawater netpens. The problems in controlling water flow make it difficult, if not impossible, to exclude pathogens in the water column from netpens. Therefore, marine pathogens are among the most important causes of diseases in seawater netpens. As salmonids are reared in fresh water before they are held in seawater, freshwater pathogens may be transferred with them to sea cages. Brown and Bruno (Chapter 4) deal with these freshwater diseases, and our emphasis is infectious diseases that are contracted after transfer to sea cages. Viral Diseases Several viruses are important pathogens of salmonid fishes, particularly during their early development in fresh water (Wolf, 1988). Viral diseases of fishes have histori- cally been of great concern to fish health managers because they can cause high mor- tality. In addition, the presence of certain viruses in a fish population causes eco- nomic hardships to fish farmers due to restrictions on transfer or sale of these fish. At least six viral diseases are of concern for pen-reared salmon: these include infectious haematopoietic necrosis (IHN), infectious pancreatic necrosis (IPN), salmon pancreas disease (SPD), infectious salmon anaemia (ISA), salmonid herpesvirus 2 infections and erythrocytic inclusion body syndrome (EIBS). The erythrocytic necrosis virus has the potential to infect salmon in seawater, ©CAB International 2002. Diseases and Disorders of Finfish in Cage Culture (eds P.T.K. Woo, D.W. Bruno and L.H.S. Lim) 61 * Current address: Center for Fish Disease Research, Department of Microbiology, Oregon State University, Corvallis, OR 97331, USA. 71
Transcript of 3 Infectious Diseases of Coldwater Fish in Marine and ... · Clinical signs of IPN in post-smolt...

3 Infectious Diseases of Coldwater Fishin Marine and Brackish Water
Michael L. Kent1,* and Trygve T. Poppe21Department of Fisheries and Oceans, Biological Sciences Branch, Pacific BiologicalStation, Nanaimo, British Columbia V9R 5K6, Canada; 2Department of Morphology,
Genetics and Aquatic Biology, The Norwegian School of Veterinary Science,PO Box 8196 Dep., N-0033 Oslo, Norway
Introduction
Salmonids are the primary fishes reared incold seawater netpens. This component ofthe industry produces approximately500,000 t year−1 on a worldwide basis.The principle species reared in netpensare Atlantic salmon (Salmo salar), cohosalmon (Oncorhynchus kisutch), chinooksalmon (Oncorhynchus tshawytscha) andrainbow trout (Oncorhynchus mykiss).Additional species include minor produc-tion of Arctic char (Salvelinus alpinus),Atlantic cod (Gadus morhua), haddock(Melanogrammus aeglefinus), Atlantichalibut (Hippoglossus hippoglossus) andAtlantic wolffish (Anarhichas lupus). Thepurpose of this chapter is to review the mostimportant infectious diseases affecting fishreared in cold seawater netpens.
The problems in controlling waterflow make it difficult, if not impossible, toexclude pathogens in the water column fromnetpens. Therefore, marine pathogens areamong the most important causes of diseasesin seawater netpens. As salmonids arereared in fresh water before they are heldin seawater, freshwater pathogens may be
transferred with them to sea cages. Brownand Bruno (Chapter 4) deal with thesefreshwater diseases, and our emphasis isinfectious diseases that are contracted aftertransfer to sea cages.
Viral Diseases
Several viruses are important pathogens ofsalmonid fishes, particularly during theirearly development in fresh water (Wolf,1988). Viral diseases of fishes have histori-cally been of great concern to fish healthmanagers because they can cause high mor-tality. In addition, the presence of certainviruses in a fish population causes eco-nomic hardships to fish farmers due torestrictions on transfer or sale of these fish.At least six viral diseases are of concern forpen-reared salmon: these include infectioushaematopoietic necrosis (IHN), infectiouspancreatic necrosis (IPN), salmon pancreasdisease (SPD), infectious salmon anaemia(ISA), salmonid herpesvirus 2 infectionsand erythrocytic inclusion body syndrome(EIBS). The erythrocytic necrosis virus hasthe potential to infect salmon in seawater,
©CAB International 2002. Diseases and Disorders of Finfish in Cage Culture(eds P.T.K. Woo, D.W. Bruno and L.H.S. Lim) 61
* Current address: Center for Fish Disease Research, Department of Microbiology, Oregon State University,Corvallis, OR 97331, USA.
71Z:\Customer\CABI\A4337 - Woo\A4419 - Woo Vouchers.vpTuesday, November 12, 2002 3:08:47 PM
Color profile: DisabledComposite Default screen

but has yet to be recognized as a seriousproblem. At least one form of plasmacytoidleukaemia is associated with a retrovirus,but this disease is described under thesection on Nucleospora salmonis. Cardiacmyopathy syndrome (CMS) has recentlybeen associated with a nodavirus, and isthus covered in this section. IHN, IPN andEIBS are also important diseases in freshwater, and are dealt with in more depthin Chapter 4. However, the manifestationsof IHN and IPN as they occur in seawaterare described. Furthermore, viruses havecaused disease in farmed Atlantic halibutand turbot larvae, resulting in considerablelosses (Bloch et al., 1991; Grotmol et al.,1995; Wood et al., 1996). However, to date,specific viral diseases of non-salmonidsreared in coldwater sea cages have not beenidentified as serious problems. Vacuolatingencephalopathy and retinopathy (VER) is adisease primarily of larval or juvenilemarine fishes, but may be carried intocage-cultured flatfish. This nodavirus isdealt with in Chapter 9.
Infectious haematopoietic necrosis in netpens
The first confirmed report of IHN in pen-reared Atlantic salmon occurred in 1992 inBritish Columbia (Armstrong et al., 1993;Traxler et al., 1993). Since this initial find-ing, IHN has been reported at many netpenfarm sites and has become a major diseaseconcern on Atlantic salmon farms in theQuadra Island region of British Columbia.The existence or establishment of marinehosts or reservoirs of IHN virus (IHNV) thatmay serve as sources of the virus at seacage sites, and the reports of virus innon-salmonids around netpens during anoutbreak is of concern (Traxler and Richard,1996). In addition, IHNV has been found inPacific herring (Clupea herningus pallasi)collected well away from infected farms(Kent et al., 1998).
Clinical signs of IHN in Atlantic salmonin netpens are often similar to those seenin infected salmonids in fresh water (seeChapter 4). However, because affected fish
are much larger than their counterparts infresh water, IHN in Atlantic salmon reared inseawater is usually chronic. Infected salmonalso often exhibit frank haemorrhages in thevisceral cavity. As with the freshwater formof the disease, IHN is best managed by avoid-ance. There is circumstantial evidence tosuggest a marine reservoir is the primarysource of the infection for outbreaks in sea-water netpens. If this were the case, thenavoidance of the infection in netpens wouldbe very difficult. Marine-phase chinooksalmon may harbour the virus for severalmonths with no signs of the disease, and thevirus has been found in healthy chinookreared at netpen farms that have experiencedIHN outbreaks in Atlantic salmon (St-Hilaireet al., 2001). Therefore, chinook salmon mayact as a subclinical reservoir for the viruswhen they are reared with Atlantic salmon.
Infectious pancreatic necrosis
This is a freshwater viral disease (seeChapter 4) that causes mortality in sea cageculture. The infection is prevalent inpen-reared Atlantic salmon in Norway(Krogsrud et al., 1989). For many years,the virus was extremely widespread inNorwegian sea sites (Melby et al., 1991)without causing clinical disease. However,in recent years clinical IPN has impactedon sea-farmed Atlantic salmon culture andis considered as an emerging problem inChapter 9. Most outbreaks occur from afew weeks up to a couple of months aftertransfer to seawater (Jarp et al., 1994), butoutbreaks up to 1 year after transfer to sea-water may also occur (Smail et al., 1992,1995). IPNV may be the most importantinfectious disease of farmed fish in Norway,accounting for losses of approximatelyNkr 400 million year−1 (Christie, 1996).In Scotland, significant mortality has beenassociated with the infection, particularlyin combination with SPD.
Clinical signs of IPN in post-smoltAtlantic salmon may be minimal, butsome fish stop feeding and show nervousdistress. The most significant losses may
62 M.L. Kent and T.T. Poppe
72Z:\Customer\CABI\A4337 - Woo\A4419 - Woo Vouchers.vpTuesday, November 05, 2002 11:34:04 AM
Color profile: DisabledComposite Default screen

sometimes be attributed to the long-termeffects of reduced or completely ceased feed-ing. Fish show hyperaemia and petechialhaemorrhage in the visceral fat and in thepyloric caeca.
An epizootiological study of IPN inpost-smolts has shown that the risk ofclinical disease was related to the mixingof smolts from several suppliers at the samesea site (Jarp et al., 1994). A way to avoidthis would therefore be to buy smolts fromas few producers as possible. Smolts withno history of IPN in fresh water, but withspecific humoral immunity against IPNVprior to smoltification, were protectedagainst clinical IPN for up to 4 months aftertransfer to seawater (Jarp et al., 1996).
Because IPNV can be vertically trans-mitted and infected fish can excrete virusfor the rest of their life, the only effectivecontrol method is avoidance. The use ofIPNV-free broodstock, rearing progeny invirus-free water and restricting the move-ment of fish are measures that can reducethe spread of IPNV. A multivalent vaccine,which includes Escherichia coli-expressedIPNV proteins, protects pre-smolt Atlanticsalmon against natural exposure to IPN(Christie, 1996). This vaccine is nowlicensed in Norway, and results from the1996 season were promising, as mortalitiesdue to IPN were reduced considerably.
Salmon pancreas disease
SPD of pen-reared Atlantic salmon is animportant disease in Scotland, Ireland andNorway (Munro et al., 1984; Ferguson et al.,1986; McVicar, 1987; Menzies et al., 1996).Hisological changes consistent with SPDhave been observed in pen-reared Atlanticsalmon in Washington State (Kent and
Elston, 1987), and rarely in BritishColumbia.
A toga-like virus has been isolated fromfish with SPD (Nelson et al., 1995), andMcLoughlin et al. (1996) experimentallyreproduced the disease with the virus.Therefore, the evidence is essentially con-clusive that the cause of pancreas disease isthis virus, referred to as salmon pancreasdisease virus (SPDV). SPD is contracted aftersmolts are transferred to seawater. Althoughnot specifically associated with fresh water(McVicar, 1987), a similar toga-like virus hasbeen isolated from rainbow trout (Boucheret al., 1994).
Fish usually exhibit clinical signs ofSPD about 6–12 weeks after introduction tonetpens, but fish that have been in pens foras long as 2 years may be affected (McVicar,1987). Mortality associated with the diseaseis low. Nevertheless, surviving fish oftenshow poor growth and are more susceptibleto other diseases (McVicar and Munro,1987).
Clinical and gross pathological changes.Affected fish are anorexic, dark in colour,lethargic, and usually at the sides of cagesand near the surface. Fish with SPD areusually emaciated (Fig. 3.1). Internal signsinclude haemorrhage in the pancreatictissue and fat between the pyloric caeca, orthe tissue between the pyloric caeca may beseverely atrophied.
Diagnosis. Haemorrhage in tissues associ-ated with the pyloric caeca in emaciatedAtlantic salmon smolts, along with theabsence of other infectious agents (e.g. IHNor IPN viruses, Aeromonas salmonicida orVibrio spp.), is indicative of SPD. Confirma-tion of the disease is based on histologicalchanges or by isolation of SPDV from
Infectious Diseases of Coldwater Fish in Salt Water 63
Fig. 3.1. Emaciated Atlantic salmon post-smolts with pancreas disease.
73Z:\Customer\CABI\A4337 - Woo\A4419 - Woo Vouchers.vpTuesday, November 05, 2002 11:34:04 AM
Color profile: DisabledComposite Default screen

affected fish. Isolation of the virus can beachieved by co-cultivation of kidney tissueson CHSE-214 cells at 15°C, and cultures areblind passed after 28 days (Nelson et al.,1995) where the cytopathic effect (CPE) maybe observed after about 10 days.
Control and treatment. No treatment isknown for pancreas disease. Reports fromScotland indicate that reducing stressors(e.g. transport and handling) during theacute phase of the disease may enhancerecovery. In addition, some farmers havereported that keeping fish on a smallerpellet size reduces anorexia and the overallmortality associated with SPD. Recoveredfish exhibit strong protection against rein-fection (Houghton, 1994), indicating that avaccine could be produced against the virus.
Infectious salmon anaemia
ISA was first observed in southwest Norwayin 1984. During the following 10 years, thedisease spread to most fish-farming areasalong the coast, but only seawater farmsand freshwater farms that use some sea-water have experienced natural outbreaks(Thorud and Djupvik, 1988). However, ISAcan be experimentally transmitted to fishin fresh water and therefore might pose athreat to these stocks. A virus typical oforthomyxoviruses has been shown to be thecause of ISA (Dannevig et al., 1995). Thevirus is shed from infected carriers beforethey develop clinical signs of the diseasethrough skin mucus, urine and faeces. Earlycolonization of the causative virus occursin the pillar cells of the gills and the endo-cardium, indicating that the gills are themost likely port of entry (Totland et al.,1996). Natural outbreaks in fish farms arerestricted to Atlantic salmon, but othersalmonids may harbour the virus andmay act as reservoirs (Nylund and Jakob-sen, 1995). ISA virus has been considereda Norwegian problem, but recently hasbeen found in Scotland, Canada andthe USA. ISA as an emerging disease isreviewed in Chapter 9.
Clinical signs and gross pathology. Mostclinical cases occur during rapid tempera-ture increases in the spring, but outbreaksmay also occur in the late autumn. Fishare anorexic, lethargic, and tend to stay atthe bottom or rest near the edges of cages.Mortality may vary from 15 to 100%. Out-breaks are usually chronic (lasting severalmonths), but are more acute if water tem-peratures are above 10–12°C. Affected fishshow a distended abdomen, exophthalmos,oedema and haemorrhage of the skin. Fishare anaemic, and the gills and heart maybe extremely pale. The visceral organs arecongested and haemorrhage occurs inthe perivisceral fat. In some cases theliver may appear extremely congestedand almost black in colour (Evensen et al.,1991b).
Diagnosis. A diagnosis is based uponcharacteristic gross pathology and lightmicroscopy, anaemia and absence of patho-genic bacteria. Supporting diagnosis of thevirus is achieved by culture on the salmonhead kidney cell line (SHK-1) (Danneviget al., 1995), a polymerase chain reaction(PCR) test (Mjaaland et al., 1997) oran enzyme-linked immunosorbent assay(ELISA) test.
Control and treatment. To control ISA,Norway has implemented strategies, includ-ing mandatory health control in smoltfarms, disinfection of processing waterfrom slaughtering facilities, separation ofdifferent year classes, isolation of infectedsites from unaffected sites and fallowing ofsites after infected stocks are removed. InCanada, commercially developed vaccinesto control ISA virus have been deployed.Recently, Jones et al. (1999) demonstrated areduction in vaccinate mortality (P < 0.01)using viral antigen emulsified in mineraloil in Atlantic salmon parr. Although trailsand licensing may be incomplete, underCanadian Regulatory Guidelines the releaseof some vaccine for use in the industry ispossible. Under current UK and Norwegianlegislation there is a restriction on the use ofvaccines for ISA.
64 M.L. Kent and T.T. Poppe
74Z:\Customer\CABI\A4337 - Woo\A4419 - Woo Vouchers.vpTuesday, November 05, 2002 11:34:05 AM
Color profile: DisabledComposite Default screen

Salmonid herpesvirus 2 infections
Several members of the family Herpesviri-dae are recognized as fish pathogens (Wolf,1988). In Japan, a herpesvirus type 2 (SH-2)infection has caused up to 30% mortalityin pen-reared coho salmon (Kumagai et al.,1994). The disease affects fish from lessthan 100 g to 1 kg, and epizootics usuallylast from 30 to 80 days.
Certain strains of salmon herpesvirus2 (e.g. Oncorhynchus masou virus (OMV)and yamame tumour virus (YTV)), causeliver damage in young fish in fresh water.Fish that survive the infection may laterdevelop epithelial tumours in sea cages(Kimura et al., 1981a,b; Sano et al., 1983;Kimura and Yoshimizu, 1991; Yoshimizuet al., 1995).
Clinical signs and gross pathology. Affectedfish are dark in colour, and often haveskin ulcers and erosion of the fins. The liverexhibits focal pale areas, and the intestinaltract shows erythema. Surface tumoursappear as whitish papillomatous massesaround the mouth, eyes, fins or gills. Thesetumours may also occur in the visceralcavity (Kimura et al., 1981a,b).
Diagnosis. Focal necrosis of the liver incoho salmon reared in Japan is presumptivediagnosis for the disease. Diagnosis isachieved by isolation of the virus fromaffected livers on CHSE-214 or RTG-2 celllines. Syncytia formation occurs in the latter(Sano et al., 1983).
Control and treatment. As with other viraldiseases, the best method to control theinfection is avoidance. The occurrence ofthe disease is associated with previousinfections at freshwater hatcheries, althoughpen-to-pen transmission in seawater isnegligible (Kumagai et al., 1997). Rainbowtrout may have subclinical infections andserve as reservoir hosts. Kumagai et al.(1997) recommended the following tocontrol the infection: (i) do not rear othersalmonids with coho salmon; (ii) disinfectfacilities after out-planting stocks; (iii) avoid
smolts from contaminated hatcheries; and(iv) examine fish for virus shortly afterintroduction to seawater.
Kimura et al. (1983) reported that dailyimmersion of chum salmon in the anti-viral compound acyclovir suppressed thegrowth of OMV-associated tumours. Theyalso found that oral treatment with anotheranti-viral drug, IUdR, decreased mortalitydue to the infection. Surface tumours areoften removed manually at harvest from fishbefore they are sent to market.
Cardiomyopathy syndrome in cage culture
This chronic, progressive disease has beenobserved since 1984 in farmed Atlanticsalmon in Norway and a few cases havebeen diagnosed in the Faroe Islands (Brunoand Poppe, 1996). The cause(s) has not beendetermined, but recently Grotmol et al.(1997) reported a nodavirus-like agent inaffected heart tissue. Although transmissionexperiments have been negative, viralparticles have been observed using elec-tron microscopy and the lesions andepizootiology are consistent with a viralaetiology. The most serious losses typicallyoccur in the autumn, 12–18 months aftertransfer to seawater.
Clinical signs and gross pathology. Fish inthe terminal stages of the disease are oftenin good body condition, showing no or fewclinical signs before death. They may gooff their feed and swim sluggishly aroundfor a few days before they die. Such fishfrequently develop skin haemorrhage andoedema, exophthalmia and ascites. Typicalfindings at necropsy are fibrinous peritoni-tis, ascitic fluid and blood clots surroundingthe heart. The atrium and sinus venosusare usually dilated and may contain bloodclots. Sometimes clotted blood may also befound on the dorsocranial surface of the liver(Ferguson et al., 1990).
Diagnosis. The diagnosis is based on thecharacteristic gross and pathognomonichistopathological lesions. Characteristic
Infectious Diseases of Coldwater Fish in Salt Water 65
75Z:\Customer\CABI\A4337 - Woo\A4419 - Woo Vouchers.vpTuesday, November 05, 2002 11:34:05 AM
Color profile: DisabledComposite Default screen

lesions are found in the spongious myo-cardium of the atrium and ventricle (Aminand Trasti, 1988; Ferguson et al., 1990).These lesions are comprised of musculardegeneration, proliferation of the endo-cardial cells with macrophage infiltrationand lymphocytes subendocardially and inthe degenerated muscle. Blood clots are fre-quently found in the atrium. Focal necrosisin the hepatic parenchyma may also occur.
Diseased fish may also be diagnosed bymeans of ultrasound imaging (Sande andPoppe, 1995). CMS bears little resemblanceto other diseases, but haemopericardiummay be observed in fish dying from otherdiseases.
Control and treatment. There are differ-ences in susceptibility to CMS between fishfamilies, and selective breeding may be apossibility for controlling this infection inthe future. Fallowing of sites for a year or twobefore new fish are introduced into problemareas has reduced the problem considerably.
Bacterial Diseases
A number of bacterial diseases causeserious and recurring losses in pen-rearedsalmon and other coldwater fishes. Someimportant bacterial diseases, such as bacte-rial kidney disease (BKD), furunculosis andyersiniosis, primarily occur in fresh waterand are dealt with in Chapter 4. Importantbacterial infections on sea pens include:typical vibriosis, caused by Vibrioanguillarum and Vibrio ordalii; coldwatervibriosis or Hitra disease caused by Vibriosalmonicida; ‘winters ulcers’ caused byMoritella viscosa (Vibrio viscosus);myxobacteriosis, caused by Cytophaga–Flexibacter spp.; and salmonid rickettsialsepticaemia or piscirickettsiosis, caused byPiscirickettsia salmonis. All salmon speciesreared in netpens are susceptible to thesebacterial diseases, but some diseases aremore problematic in certain species andparticular areas. For example, chinook,coho and sockeye salmon appear to be moresusceptible to BKD than Atlantic salmon,
whereas furunculosis and myxobacteriosisrepresent an increased problem for Atlanticsalmon compared with Pacific salmonspecies. Almost all non-salmonid marinefishes are susceptible to vibriosis and furun-culosis. Typical furunculosis is causedby A. salmonicida subsp. salmonicida. Theatypical strain of A. salmonicida is usuallythe aetiological agent of furnuculosis innon-salmonid marine fishes.
Vibriosis
Vibriosis is a systemic disease that affectsmany marine fishes and invertebrates(Anderson and Conroy, 1970; Colwell andGrimes, 1984; Egidius, 1987). Frerichs andRoberts (1989) considered vibriosis to bethe most significant disease in wild andcultured marine and brackish water fishes.V. anguillarum accounts for most of theoutbreaks of vibriosis in farmed salmonworldwide, and also causes disease inAtlantic cod. V. ordalii occasionally causesdisease in salmonids reared in the PacificNorthwest and in New Zealand (Evelyn,1971; Harrell et al., 1976; Novotny, 1978;Schiewe et al., 1981; Wards et al., 1991).Diseases caused by other Vibrio spp.include coldwater vibriosis or Hitra disease,caused by V. salmonicida, and winter ulcerdisease caused by M. viscosa. The latter isconsidered as an emerging problem and isdiscussed in Chapter 9.
V. anguillarum strains show hetero-geneity in both phenotypic (Tajima et al.,1985) and serotypic (Kitao et al., 1984;Tajima et al., 1985; Sorensen and Larsen,1986) characteristics. The strains of V.anguillarum that cause vibriosis in pen-reared salmon worldwide represent onlyone or two serotypes (based on the ‘O’antigens present), which simplifies theformulation of anti-vibriosis vaccines forcontrolling the disease. Vibriosis causedby V. anguillarum usually occurs between15 and 21°C, and most outbreaks occur insmolts during their first summer in seawater.Vibriosis in the Pacific Northwest is mainlya problem of Pacific salmon (e.g. chinook
66 M.L. Kent and T.T. Poppe
76Z:\Customer\CABI\A4337 - Woo\A4419 - Woo Vouchers.vpTuesday, November 05, 2002 11:34:05 AM
Color profile: DisabledComposite Default screen

and coho), although Atlantic salmon are alsosusceptible to the infection. Interestingly,all cases of vibriosis due to V. ordalii inpen-reared salmon reported to date haveinvolved Pacific salmon in Japan and thePacific coast of the USA (Schiewe et al.,1981).
Coldwater vibriosis is a bacterialsepticaemia caused by the psychrophilicbacterium V. salmonicida. Since its firstoccurrence in farmed Atlantic salmon innorthern Norway in 1977 (Egidius et al.,1981), coldwater vibriosis has been diag-nosed in most fish-farming areas, as well asin salmon-producing countries surroundingthe North Atlantic (Bruno et al., 1986),including eastern Canada and the USA(O’Halloran and Henry, 1993). The condi-tion is also known as ‘Hitra disease’ aftersevere outbreaks occurred in the Hitra regionof Norway in the early 1980s. The disease isusually most severe at low temperaturesduring the winter months, but may occurthroughout the year. Environmental stress-ors and poor nutrition may predisposefish to coldwater vibriosis. Although thebacterium may cause disease in otherfish, such as Atlantic cod (Jøtorgensen et al.,1989), serious losses occur mainly in Atlan-tic salmon. Schroder et al. (1992) showedin experimental studies that cod are moreresistant to the bacterium than salmon.
Clinical signs and gross pathology. In smallfish, mortality caused by vibriosis may behigh and rapid, and these fish may exhibitno gross pathological changes other thandarkening and lethargy. Typical of bacterialsepticaemias, fish with vibriosis may exhibiterythema at the base of the fins, petechiaein the skin and haemorrhage on the bodysurface. Fish may also exhibit bilateralexophthalmia and frayed fins. Haemor-rhagic abscesses in the muscle are often seenin Atlantic salmon with vibriosis in Europe.Congestion and petechiae are usuallyevident in visceral organs, particularly inthe gut and liver. Large multiple coalescinghaematomas in the liver (peliosis hepatis)are often seen in vibriosis caused by V.anguillarum. Affected fish also exhibitpallor of the gills (due to anaemia) andenlargement of the spleen and kidney.
Clinical signs of coldwater vibriosismay be non-specific, but usually includelethargy and cessation of feeding. Affectedfish turn dark, exhibit exophthalmos, aswollen vent and pinpoint haemorrhagealong the belly and at the base of the pecto-ral, pelvic and anal fins. The gills are usuallypale. Internally, ascites and petechial haem-orrhage in the perivisceral fat, pyloric caeca,peritoneal surfaces, liver and swimbladderare typical findings (Figs 3.2 and 3.3). Thelatter may be filled with a blood-tinged fluid
Infectious Diseases of Coldwater Fish in Salt Water 67
Fig. 3.2. Atlantic salmon with coldwater vibriosis. Note multiple haemorrhage in the liver.
77Z:\Customer\CABI\A4337 - Woo\A4419 - Woo Vouchers.vpTuesday, November 05, 2002 11:34:06 AM
Color profile: DisabledComposite Default screen

and the liver typically has a yellowish dis-coloration. In chronic cases, skin ulceration,fin rot, a pseudomembranous peritonitis andepicarditis may also be found. The spleenusually has a colour slightly lighter than nor-mal. In cod, the pathological changes arerather diffuse and non-specific, but keratitisis frequently seen.
Diagnosis. For typical vibriosis, presump-tive diagnosis is by macroscopic examina-tion if the characteristic haematomas inthe liver are present. The causative Gram-negative bacilli are usually easy to detectin Gram-stained kidney smears. The highlymotile bacteria are also detectable in freshpreparations of blood or in wet mounts ofthe kidney or spleen, or in lesions. The othergross and clinical changes are not specific tovibriosis and are associated with a number ofbacterial or viral systemic diseases.
A diagnosis is based on culture andidentification of the causative organismfrom the kidney of infected fish. BothV. anguillarum and V. ordalii are easily cul-tured on tryptic soy agar with 1.5% NaCl oron marine agar (Difco) at room temperature.Bacterial colonies are round, raised andoff-white in colour. V. ordalii and V. salmon-icida grow more slowly than V. anguillarumand form smaller colonies. Optimum growthtemperature for V. salmonicida is from 12 to16°C and growth will occur between 0 and
22°C. In contrast, V. anguillarum and V.ordalii will grow at about 25°C. The bacteriacan be distinguished using biochemical tests(Schiewe et al., 1981; Holm et al., 1985;Tajima et al., 1985; Scalati and Kusuda,1986). API-20E test strips (Analytab Co.,Plainview, New York, USA) can be used forrapid identification of marine vibrios fromfish (Kent, 1982; Grisez et al., 1991). Thesebacteria can also be identified serologicallyusing slide agglutination tests.
With V. salmonicida, microscopicaldemonstration of the bacterium in Giemsa-stained smears or paraffin sections, or byimmunohistochemistry (Evensen et al.,1991a) is useful for locating the organism intissues. However, immunofluorescence testsapplied directly to tissues infected with V.anguillarum are not useful for rapid diagno-sis of this bacterium. Apparently salmonidtissues contain substances that block recep-tor sites on the vibrios that would normallyreact with the vibrio-specific antibodiesin the diagnostic antisera (T.P.T. Evelyn,Pacific Biological Station, Nanaimo, BritishColumbia, personal communication).
Control and treatment. Vibriosis is bestconrolled by prevention, and commercialvaccines are available. With salmonids,vaccination is best carried out on fish thathave attained immunocompetent size (atleast 5–10 g) and before they are introduced
68 M.L. Kent and T.T. Poppe
Fig. 3.3. Vibrio salmonicida infection in Atlantic salmon. Note petechial haemorrhage in the perivisceralfat between the pyloric caeca.
78Z:\Customer\CABI\A4337 - Woo\A4419 - Woo Vouchers.vpTuesday, November 05, 2002 11:34:06 AM
Color profile: DisabledComposite Default screen

to netpens. The vaccines are convenientlyadministered by immersion methods, andif applied properly, they afford excellentprotection (Evelyn, 1984, 1988). Some farmsrevaccinate fish shortly after introduction tonetpens, and results with revaccination inseawater have been promising. Revaccina-tion should, however, be conducted withcaution because the handling of fishshortly after their introduction to seawater isvery stressful. The most recent trend is toadminister the vaccine by intraperitonealinjection, usually in combination with othervaccines, e.g. with furunculosis vaccines,which are most effective when injected.Vaccines for controlling vibriosis in codare also promising for protecting against V.anguillarum (Espelid et al., 1991; Gromanet al., 1992) and V. salmonicida (Schroderet al., 1992).
Antibacterial drugs (e.g. oxytetracy-cline, potentiated sulphonamides, quino-lones and florfenicol) incorporated in feedare available for treating vibriosis. Treat-ment is usually efficacious if the infection isrecognized early when fish are still activelyfeeding, and if care is taken to select a drug towhich the pathogen is sensitive. However, insome countries not all of the drugs have beenapproved for use in fish intended for humanconsumption. Thus, control of vibriosisshould be conducted primarily through avaccination programme.
V. salmonicida is not considered to be ahighly pathogenic bacterium and significantexposure is required to infect fish. As withother diseases, optimization of the environ-ment and reduction of stressors, particularlyduring the winter months, are importantmeasures to avoid outbreaks. Multivalentvaccines protecting against furunculosis,vibriosis and coldwater vibriosis giveexcellent protection provided the vaccina-tion programmes are carried out in a propermanner (e.g. fish are vaccinated at anappropriate size and given adequate timeto develop immunity before exposure). Nev-ertheless, outbreaks do occur in properlyvaccinated fish, particularly in northernNorway. Although the bacterium occurscommonly in the water and sediments closeto cages, its numbers escalate during
outbreaks and it is therefore important toisolate diseased fish from healthy fish (Engeret al., 1989) and to remove dead and mori-bund fish from the cages.
Marine myxobacteriosis
Cytophaga and Flexibacter spp. (commonlyreferred to as ‘myxobacteria’ or ‘glidingbacteria’) are important bacterial pathogensof cultured fishes and usually causeexternal lesions in freshwater and marinespecies (Anderson and Conroy, 1969; Pachaand Ordal, 1970). In marine aquaculture,infections by Flexibacter maritimus havebeen observed in Japanese flounder(Paralichthys olivaceous) and seabreams(family Sparidae) in Japan (Masumura andWakabayashi, 1977; Hikida et al., 1979;Wakabayashi et al., 1984, 1986; Baxa et al.,1986, 1987) and Europe (Bernardet et al.,1990). In Tasmania, Handlinger et al. (1997)identified F. maritimus associated with skinand gill lesions in pen-reared Atlanticsalmon and rainbow trout. Myxobacteriathat have not been precisely identified havebeen associated with skin lesions in sea-water-reared salmonids for many years(Borg, 1960; Rucker et al., 1963; Andersonand Conroy, 1969; Wood, 1974; Sawyer,1976). These bacteria have at times beenidentified as Sporocytophaga spp. How-ever, the presence of microcysts (an impor-tant diagnostic feature of this genus) has notbeen clearly demonstrated in these isolates.
Proper taxonomic identifications havenot been conducted on many Cytophaga andFlexibacter spp. that have been associatedwith disease in marine fishes. However, todate, marine Cytophaga and Flexibacterspecies have not been transferred to Flavo-bacterium (Bernardet et al., 1996), as wasproposed for certain freshwater species. Fishhealth workers and aquaculturists usuallyrefer to these bacteria as ‘myxobacteria’.This is technically incorrect because thesebacteria belong to the order Cytophagales,and not to the order Myxobacteria. It would,therefore, be more appropriate to refer tothe marine forms using collective terms such
Infectious Diseases of Coldwater Fish in Salt Water 69
79Z:\Customer\CABI\A4337 - Woo\A4419 - Woo Vouchers.vpTuesday, November 05, 2002 11:34:07 AM
Color profile: DisabledComposite Default screen

as ‘cytophaga–flexibacter-like bacteria’ or‘gliding bacteria’. However, to remain con-sistent with the common terminology and toavoid confusion, reference is made to thesebacteria as myxobacteria in this review.
Two types of myxobacterial infectionshave been associated with high mortality inpen-reared Atlantic salmon in the PacificNorthwest; one type causes large bodyulcers, and the other causes lesions primar-ily in the mouth. Myxobacteria infectionsare also seen in pen-reared Pacific salmon,but are not usually associated with severeepizootics. In Pacific salmon, myxobacteriaare usually associated with frayed fins anderosion of the tail.
A Cytophaga sp. causes large skinlesions in Atlantic salmon smolts (Fig. 3.4)shortly after transfer to seawater (Kent et al.,1988). Very similar skin lesions associatedwith F. maritimus infections were observedin pen-reared Atlantic salmon and rainbowtrout in Tasmania (Handlinger et al., 1997).Lesions and associated mortalities usuallypeak at about 1–3 weeks after introduction,and based on our observations, the infectionsubsides after about 3–4 weeks. There
appears to be a seasonality, and fishintroduced later in the spring and summerusually exhibit fewer body lesions. Infec-tions are restricted to the skin and muscle.Fish with large lesions apparently die fromosmotic imbalance (Kent et al., 1988a).
A particularly lethal form of myxobac-teriosis occurs in the mouth of pen-rearedAtlantic salmon, referred to as ‘mouth rot’by fish farmers. Infections of the mouthand snout by myxobacteria are observedin post-smolt Atlantic salmon during theirfirst summer in seawater. The condition hasoccurred at many netpen sites in the PacificNorthwest, and is often associated withhigh mortalities (Hicks, 1989; Frelier et al.,1994). Pen-reared Arctic char have also beenafflicted with the infection. The infectionappears to begin around the teeth. It has beensuggested that the infection is initiated inperiodontal tissue that has been abraded byfeeding on spiny crustaceans such as crablarvae and Caprella spp. amphipods. Otherpotential predisposing factors suggested byfarmers that may lead to the infection are: (i)feeding on hard pellets; (ii) fish biting netsurfaces; and (iii) stress-induced lesions in
70 M.L. Kent and T.T. Poppe
Fig. 3.4. Atlantic salmon with Cytophaga-associated skin lesions. Note severe lesion with exposure ofunderlying muscle.
80Z:\Customer\CABI\A4337 - Woo\A4419 - Woo Vouchers.vpTuesday, November 05, 2002 11:34:08 AM
Color profile: DisabledComposite Default screen

the mouth. Fish farmers in British Columbia,Canada report that the condition is par-ticularly troublesome in waters with highsalinity. Based on preliminary culture analy-sis, the myxobacterium from mouth lesionsappears to be different from the myxo-bacterium causing skin lesions. A similarmyxobacterial stomatitis has been observedin wild Atlantic cod in the North Sea (Hilgeret al., 1991).
Clinical signs and gross pathology. Skinlesions are large, white patches on thecaudal peduncle and the posterior region ofthe flanks when the dermis is intact. Fishwith more severe lesions have areas of theskin completely destroyed and the under-lying muscle exposed (Fig. 3.4). Fish withmouth myxobacteriosis are often lethargic,emaciated and anorexic, and some affectedfish may exhibit flashing or head shaking.Early in the infection, examination of themouth reveals focal, yellow bacterial matsaround the palate and teeth, including thevomer. The lesions may be single, but theopposing surface is often affected (Frelieret al., 1994). As the disease progresses,affected fish show multiple ulcers in themouth with large bacterial mats overlyingthe lesions. The lesions may extend to thebranchial arches and proximal oesophagus,and the lower and upper jaw may be com-pletely eroded in severe cases. Severelyaffected fish do not feed and hence thestomach is devoid of food.
Diagnosis. Diagnosis of myxobacterialinfections can usually be accomplishedby observing large numbers of filamentousbacteria in wet-mount preparations fromthe lesions (Fig. 3.5). Isolation of the bacteriacan be accomplished by culture on eitherCytophaga medium made with 50% sterileseawater or marine agar (Difco). Isolationof myxobacteria in pure culture may bedifficult from skin lesions due to contam-ination with other faster-growing bacteria(e.g. vibrios). However, the lesions usuallycontain large numbers of the myxobacteriaand serial dilutions of affected tissue insterile 50% seawater facilitates the isolationof the myxobacteria in pure culture.Reichenbach (1988) has described thegeneral characteristics of Cytophaga andFlexibacter spp.
Treatment and control. External treatmentswith antibiotics are often used to controlmyxobacterial infections in fresh water,but such treatments are not usuallypractical in seawater netpens. Thesebacterial infections are often initiated inthe skin where there are abrasions. Physicaltrauma during transport of smolts mayallow the bacteria to establish an infection.According to fish farmers, improved trans-port techniques and careful handling offish greatly reduces the prevalence of thedisease. There are unconfirmed reports thattreating fish with oxytetracycline can con-trol mouth rot.
Infectious Diseases of Coldwater Fish in Salt Water 71
Fig. 3.5. Cytophaga bacteria in wet mount of skin lesions showing ‘myxobacteria’. Bar, 5 µm.
81Z:\Customer\CABI\A4337 - Woo\A4419 - Woo Vouchers.vpTuesday, November 05, 2002 11:34:08 AM
Color profile: DisabledComposite Default screen

Salmonid rickettsial septicaemia
Piscirickettsia salmonis causes a severesepticaemia in pen-reared salmon, par-ticularly in Chile (Branson and Neito Diaz-Munoz, 1991; Cvitanich et al., 1991; Garcéset al., 1991; Fryer et al., 1992; Almendrasand Carmeu Fuentealba, 1997). In BritishColumbia, the disease was first observedin seawater-reared pink salmon (Oncorhyn-chus gorbuscha), held for experimentalpurposes, at the Pacific Biological Stationon Vancouver Island in 1970. Morerecently, infections of Atlantic salmonwith rickettsia-like organisms have beenreported from Norway (Olsen et al., 1997),Ireland (Rodger and Drinan, 1993) andScotland (Grant et al., 1996). A PCR methoddeveloped by Mauel et al. (1996) for detect-ing and identifying the pathogen showedthat the isolates from Norway, Ireland,Canada and Chile were all P. salmonis; itwas clear that at least two variants of thepathogen occurred in Chile. House et al.(1998) showed that the strain from Chilewas more pathogenic than common strainsfrom British Columbia and Norway.
In British Columbia, the infection isusually coincidental with other infectiousdiseases (e.g. bacterial kidney disease) in thepopulation, but may occasionally causeepizootics in which it is the primary causeof mortality. In contrast, piscirickettsiosisis the most important infectious diseaseof pen-reared salmonids in Chile, where itcaused losses of around US$48 million in1995. In Chile, the disease was first recog-nized as a serious problem in coho salmon,but it now is also common in both rainbowtrout and Atlantic salmon. Several outbreaksof the disease may occur in the same popula-tion of fish during their seawater grow-outperiod, particularly with coho salmon.
Information to date on the epizootiologyof the organism suggests that it is normallyacquired in seawater from a marine source.However, a marine reservoir has yet to beidentified, although certain salmon ecto-parasites may be involved in the trans-mission, perhaps serving as vectors (Garcéset al., 1994). Furthermore, Cvitanich et al.(1991) found evidence of the organism in
crustaceans and molluscs around netpens,based on histology and serology. Neverthe-less, P. salmonis survives well in seawater(Almendras, 1996) and is easily transmitteddirectly from fish to fish.
The disease may also occur in brackishwater, and the infection has recently beenreported in rainbow trout and coho salmonheld in fresh water (Bravo, 1994; Cvitanichet al., 1995; Gaggaro et al., 1995). However,P. salmonis does not survive in fresh wateraway from the host (Lannan and Fryer,1994). Bustos et al. (1994) conducted fieldtrials that suggested that vertical transmis-sion may occur naturally, and Larenas et al.(1996) detected the infection in 10% of fertil-ized ova from infected fish. This may explainits occurrence in fresh water. However, thepoor survivability of the organism in freshwater may explain the rarity of the infectionbefore fish are introduced to seawater.
Clinical signs and gross pathology. Clinicaland gross pathological changes associatedwith P. salmonis infections have been out-lined by Cvitanich et al. (1991), Branson andNieto Diaz-Munoz (1991) and Brocklebanket al. (1992). Affected fish are lethargic,anorexic, exhibit pallor of the gills due toanaemia, are dark in colour and may swimnear the surface. There are marked differ-ences in clinical signs between salmonidspecies. For example, infected rainbow troutoften do not accumulate near the surfacebefore they die. In Atlantic and coho salmonthe nervous system is often affected, withflashing and side swimming being commonin the former. Ulceration often occurs on theskin with coho salmon and rainbow trout,whereas this is rare with Atlantic salmon inChile. However, in Norway, Atlantic salmonwith the disease occasionally show skinlesions, e.g. raised nodules or white spots(Olsen et al., 1997).
The liver of affected fish may have large,whitish or yellow, multifocal, crater-likelesions or nodules (Fig. 3.6). These lesionsoften rupture, resulting in shallow crater-like cavities in the liver. Fish may haveascites, an enlarged spleen and a grey,enlarged kidney. The spleen is extremelyenlarged in infected pink salmon. Pallor and
72 M.L. Kent and T.T. Poppe
82Z:\Customer\CABI\A4337 - Woo\A4419 - Woo Vouchers.vpTuesday, November 05, 2002 11:34:08 AM
Color profile: DisabledComposite Default screen

petechiae are observed in the visceral organsand muscle, and a whitish pseudomembranemay cover the heart.
Diagnosis. Presumptive diagnosis can beachieved by observing the distinctivecrater-like lesions and nodules in the liver,but they may not occur in many infectedfish. Definitive diagnosis can be achieved byobserving the organism within phagocyticcells in liver or kidney imprints stainedwith Giemsa, Gram or methylene blue stains(Fig. 3.7A), or in macrophages in tissue sec-tions (Fig. 3.7B) along with the distinctivehistological changes described above.Acridine orange-stained tissue smears arealso useful for demonstrating the organism
(Lannan and Fryer, 1991). Further confirma-tion includes the isolation of the organism inculture using CHSE-214 cells (Fryer et al.,1990; Cvitanich et al., 1991), an indirectfluorescent-antibody test (Lannan et al.,1991) or using specific primers with PCR(Mauel et al., 1996). A commercial ELISAtest for P. salmonis was developed byMicrotek Ltd-Bayer (Sidney, British Colum-bia) and used extensively by Chilean farmersin brood stock segregation programmes.
Control and treatment. Various antibiotics,such as oxolinic acid, flumequine andoxytetracycline, have been used to treatthe infection, often with limited success. Inextreme cases some Chilean farmers have
Infectious Diseases of Coldwater Fish in Salt Water 73
Fig. 3.6. Piscirickettsiosis in salmon. Multiple, white, crater-like lesions in the liver (courtesy of P. Bustos).
Fig. 3.7. Piscirickettsia salmonis. (A) Gram-stained kidney imprint. (B) Organisms in macrophages fromkidney section, stained with H&E. Bar, 10 µm.
83Z:\Customer\CABI\A4337 - Woo\A4419 - Woo Vouchers.vpTuesday, November 12, 2002 3:09:08 PM
Color profile: DisabledComposite Default screen

had success with employing injectable treat-ments with fluoroquinolones. The micro-organism is intracellular, and this probablycontributes to the difficulties of treatingthe disease with antibiotics. Although therole of vertical transmission is unknown,the techniques used for preventing verticaltransmission of bacterial kidney disease (e.g.brood stock screening) are being employedin Chile with some success.
Protozoa and Myxozoa
Protozoans and myxosporeans (phylumMyxozoa) are some of the most importantpathogens of cage-reared fishes. For exam-ple, the amoeba Paramoeba pemaquidensisis an important gill pathogen in cage-reared salmon in Australia, and theflagellate Ichthyobodo (= Costia) spp.(see Chapter 4) and trichodinid ciliatesinfect gill surfaces of salmon and somemarine fishes. Systemic infections byCryptobia salmositica (see Chapter 9) and adiplomonad flagellate similar to Hexamitasalmonis have caused disease in chinooksalmon in British Columbia. Anotherdiplomonad (Spironucleus barkhanus)has caused extra-intestinal infections inAtlantic salmon in Norway. Four myxo-sporeans (Parvicapsula sp., M. aeglefini,Kudoa thyrsites and Chloromyxumtruttae) and three microsporidians (Lomasalmonae, N. salmonis and Microsporidiumcerebralis) infect internal organs or gills.Some protozoan infections are contractedin fresh water (e.g. Cryptobia, Ichthyobodo)and are dealt with in Chapter 4. In the pres-ent chapter, the most important marine pro-tozoan and myxosporean parasites affectingcoldwater marine netpen culture, i.e.Paramoeba sp., K. thyrsites, extraintestinaldiplomonads, L. salmonae and N. salmonis,are discussed.
Paramoeba pemaquidensis gill infections
Paramoeba sp. (Sarcomastigophora: Para-moebidae) has caused devastating losses in
pen-reared rainbow trout and Atlanticsalmon in Tasmania (Roubal et al., 1989;Munday et al., 1993) and Europe (Rodgerand McArdle, 1996). A similar (if not identi-cal) amoeba (identified as P. pemaquiden-sis) was associated with severe gill diseasein coho salmon reared in netpens in Wash-ington State and land-based seawater tanksin California (Kent et al., 1988b). Thisspecies is an opportunistic pathogen that isnormally free-living in seawater. Intensityand prevalence of the amoeba on fish gillsvaried from year to year, with infectionsbeing most prevalent in the late summerand autumn. The exact environmentalconditions or health status of the fish thatallow the organism to proliferate on fishgills are unknown. Presumably fish alreadycompromised by other diseases are moresusceptible to the infection, and in Wash-ington State many of the infected fishhad pre-existing diseases or smoltificationproblems (Kent et al., 1988b).
Clinical signs and gross pathology. Heavi lyinfected fish are lethargic, accumulate atthe surface and have flared opercula. Exces-sive mucus is often observed on heavilyinfected gills. Focal, whitish patches may beobserved on heavily infected fish.
Diagnosis. Paramoebiasis of salmon is diag-nosed by the detection of large numbers ofthe amoebae on the gills. The organisms arebest identified in fresh wet-mount prepara-tions of the gills. Floating and transitionalforms of the amoeba on the gills are20–30 µm in diameter and have severaldigitiform pseudopodia. In wet mounts,amoebae will attach to the slide after aboutan hour, resulting in a locomotive formmeasuring about 20 × 25 µm. Paramoebaspp. possess a unique organelle, called aparasome or Nebenkörper, which is adjacentto the nucleus (Fig. 3.8). The parasome canbe observed in wet mounts of locomotiveforms, and is readily visible with FeulgenDNA stains. Amoebae can also be identifiedon gill surfaces in histological preparations,but many detach during processing. Theamoebae can also be identified with specific
74 M.L. Kent and T.T. Poppe
84Z:\Customer\CABI\A4337 - Woo\A4419 - Woo Vouchers.vpTuesday, November 05, 2002 11:34:10 AM
Color profile: DisabledComposite Default screen

polyclonal antibodies in tissue sections orimprints (Howard and Carson, 1993a).
Treatment and control. Most compoundstypically used as external treatments (e.g.formalin, chelated copper, diquat, malachitegreen and chloramine T) are not effectiveagainst the organism (Munday et al., 1993),but the amoeba is readily eradicated fromfish gills with freshwater bath treatments.Kent et al. (1988b) confirmed that the para-site survives poorly in low salinity water.Cameron (1993) reported that reducing sea-water concentrations to 4 ppt was neededfor effective treatment. Reducing the salinityhas been effective for eradicating infectionsin fish held in land-based tanks, but thistreatment is usually difficult to apply andimpractical in netpens. Hydrogen peroxidebath treatments at concentrations between200 and 400 ppm are moderately effectiveat controlling the infection (Cameron, 1993).Howard and Carson (1993b) reported that100 ppm hydrogen peroxide for 2 h killedall the amoeba. However, Cameron (1994)found that hydrogen peroxide did controlthe infection in field situations, even whenused at 300 ppm. Hydrogen peroxide treat-ments should be applied with cautionbecause of potential toxic effects to thefish, particularly at higher temperatures
(Cameron, 1993; Johnson et al., 1993; Brunoand Raynard, 1994).
Diplomonad flagellates
Extra-intestinal infections by diplomonadflagellates have cause disease in caged-reared salmon in Norway and BritishColumbia. One outbreak of a severe sys-temic infection by a diplomonad flagel-late (family Hexamitidae) resembling H.salmonis caused close to 50% mortality inchinook at one netpen site in the Secheltarea, British Columbia (Kent et al., 1992).The fish were introduced to seawater in thespring of 1990 and showed high mortalitystarting in September 1991. Interestingly,about the same time, extra-intestinal infec-tions by a similar parasite were reported inpost-smolt to adult Atlantic salmon rearedat netpen farms in northern Norway (Moet al., 1990; Poppe et al., 1992).
Whereas gut infections in fish bydiplomonads are common (Woo andPoynton, 1995), systemic infections bydiplomonad parasites in fish are rare. Onlyone other report of such infections is known,and Ferguson and Moccia (1980) reporteda similar disease in Siamese fighting fish(Beta splendens). Although the flagellates
Infectious Diseases of Coldwater Fish in Salt Water 75
Fig. 3.8. Paramoeba pemaquidensis from coho salmon gill. The parasome (arrowhead) is adjacent to thenucleus. Bar, 10 µm.
85Z:\Customer\CABI\A4337 - Woo\A4419 - Woo Vouchers.vpTuesday, November 05, 2002 11:34:11 AM
Color profile: DisabledComposite Default screen

observed in pen-reared salmon are morpho-logically indistinguishable from the rela-tively non-pathogenic H. salmonis thatinfects the intestinal tract of salmonids infresh water, they probably represent new,highly invasive strains or species. Sterudet al. (1997) recently named the organismfrom pen-reared Atlantic salmon and Arcticgrayling (Thymallus arcticus) from freshwater as S. barkhanus.
Water-borne exposure of the fish toinfected blood and viscera, or cohabitationwith infected fish in either fresh or seawater(Kent et al., 1992) can reproduce the sys-temic disease in chinook salmon. However,the parasite of Atlantic salmon from Norwaycould not be transmitted to healthy fish bycohabitation.
Clinical and gross pathology. In chinooksalmon from British Columbia, infected fishappeared normal except that some fish had adistended abdomen. The gills were pale dueto anaemia. The hallmark gross pathologicalchange of the disease is an extremelyenlarged liver. In some fish, the liver wasalso mottled and had petechial haemorrhageand whitish, friable areas. Affected fishconsistently had serosanguinous ascites andblood clots in the visceral cavity. The spleenand kidney were moderately enlarged, andpetechiae occurred throughout the skeletalmuscle (Kent et al., 1992).
In Atlantic salmon, the infection dif-fered in that the parasite caused large,multifocal, white, lesions in the muscula-ture, liver, spleen and kidney (Poppe et al.,1992). Yellow or white cysts filled with theparasite were also in the fins, and infectedfish often had exophthalmia (Poppe and Mo,1993).
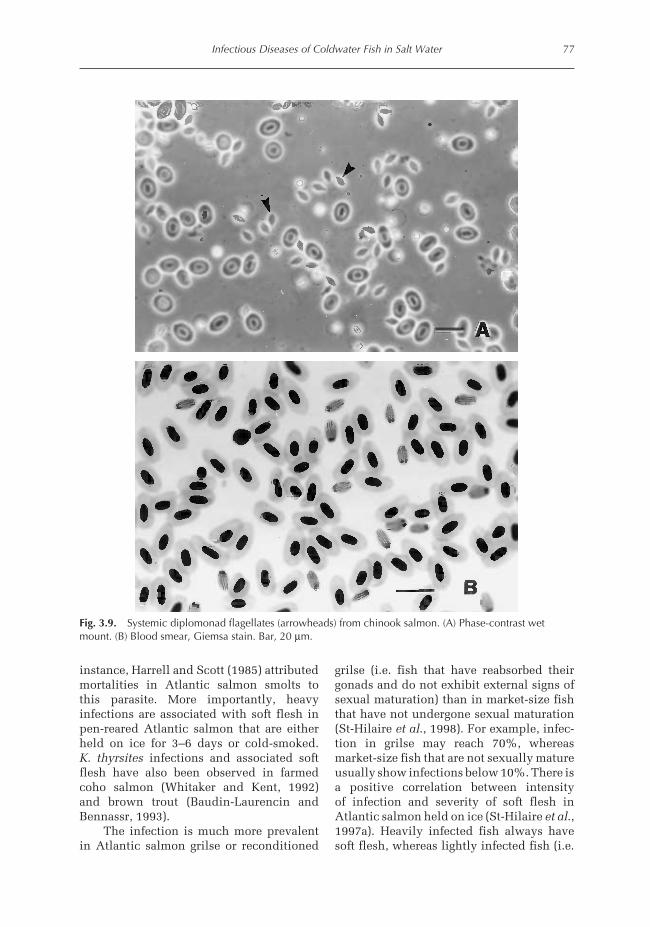
Diagnosis. The infection is identified bywet-mount preparations or Giemsa-stainedimprints of the gut or other visceral organs.Because the parasite is highly motile, it maybe easier to identify the parasites in wet-mount examination. Wet-mount prepara-tions of the visceral organs reveal massivenumbers of flagellates that are 10 × 5 µm(Fig. 3.9A). The parasites are also readily
detected in DifQuick or Giemsa-stainedimprints (Fig. 3.9B), where they appear asdark-staining, oval bodies with two clearbands, representing the flagella pocket, run-ning the length of the organism. The twonuclei at the anterior end of the parasite maybe visible.
Control and treatment. Several drugs, mostof which are added to the diet, have beenrecommended for the control of H. salmonisinfections in the gut of salmonids (Yasutakeet al., 1961; McElwain and Post, 1968;Hoffman and Meyer, 1974; Becker, 1977).However, presently, none of these com-pounds is approved for treatment of foodfish in Canada or the USA.
Although it has been determined thatthe disease is transmissible in seawater, it isnot established whether salmon contractedthe infection in seawater, or were subclini-cally infected when they were transferred tonetpens. At present, the best guess is thatinfections in both Norway and Canada arecontracted in seawater. In addition, Poppeand Mo (1993) suggested that fish couldbecome infected by exposure to untreatedwater from fish processing plants. An under-standing of the source of the infection wouldbe helpful for implementing effective con-trol strategies.
Kudoa thyrsites (Myxozoa)
Myxosporeans of the genus Kudoa andrelated genera infect the muscle of manymarine fishes, and heavy infections cancause unsightly white cysts or soft texturein fillets (Kabata and Whitaker, 1981;Patashnik et al., 1982). These parasites canlower the market value of the infectedfish, although they seldom cause morbidity.K. thyrsites is a cosmopolitan parasite thatinfects many species of marine fishes(Whitaker et al., 1994). Infections inpen-reared Atlantic salmon have beenreported from the Pacific Northwest(Harrell and Scott, 1985; Whitaker andKent, 1991), Spain (Barja and Toranzo,1993) and Ireland (Palmer, 1994). In one
76 M.L. Kent and T.T. Poppe
86Z:\Customer\CABI\A4337 - Woo\A4419 - Woo Vouchers.vpTuesday, November 05, 2002 11:34:11 AM
Color profile: DisabledComposite Default screen
instance, Harrell and Scott (1985) attributedmortalities in Atlantic salmon smolts tothis parasite. More importantly, heavyinfections are associated with soft flesh inpen-reared Atlantic salmon that are eitherheld on ice for 3–6 days or cold-smoked.K. thyrsites infections and associated softflesh have also been observed in farmedcoho salmon (Whitaker and Kent, 1992)and brown trout (Baudin-Laurencin andBennassr, 1993).
The infection is much more prevalentin Atlantic salmon grilse or reconditioned
grilse (i.e. fish that have reabsorbed theirgonads and do not exhibit external signs ofsexual maturation) than in market-size fishthat have not undergone sexual maturation(St-Hilaire et al., 1998). For example, infec-tion in grilse may reach 70%, whereasmarket-size fish that are not sexually matureusually show infections below 10%. There isa positive correlation between intensityof infection and severity of soft flesh inAtlantic salmon held on ice (St-Hilaire et al.,1997a). Heavily infected fish always havesoft flesh, whereas lightly infected fish (i.e.
Infectious Diseases of Coldwater Fish in Salt Water 77
Fig. 3.9. Systemic diplomonad flagellates (arrowheads) from chinook salmon. (A) Phase-contrast wetmount. (B) Blood smear, Giemsa stain. Bar, 20 µm.
87Z:\Customer\CABI\A4337 - Woo\A4419 - Woo Vouchers.vpTuesday, November 05, 2002 11:34:13 AM
Color profile: DisabledComposite Default screen

fewer than 20,000 spores g−1) usually do nothave the condition. Soft flesh is not noticedon the processing line, and only becomesapparent after fish are held for about 3–6days on ice or when fillets are smoked. Inan investigation of K. thyrsites infections inPacific hake (Merluccius productus), theflesh softening was apparently caused by aproteolytic enzyme produced by the parasite(Tsuyuki et al., 1982). This enzyme remainsactive below 70°C. Therefore, tissue break-down will continue through cold-smokingprocesses, which are normally conducted atabout 50°C or less. Seymour et al. (1994)suggested that the flesh degradation is dueto cathepsin L from the host inflammatoryresponse to the parasite, instead of aproteolytic enzyme from the parasite.
Very little is known about developmentand transmission of K. thyrsites in fish. Ittakes about 5–6 months after infection beforespores are detected in the flesh and a highprevalence occurs in post-smolts (Moranet al., 1999a). As the infection progresses inAtlantic salmon, pseudocysts in the musclefibres enlarge and ultimately rupture. A pro-minent inflammatory response is associatedwith ruptured pseudocysts, and fish elimi-nate the detectable infections after abouta year in seawater (Moran et al., 1999a).It is not known if the high prevalence ofthe infection in grilse is due to reinfection,or proliferation of a cryptic infection thatoriginally occurred shortly after fish were
transferred to seawater. An infectious stageof the parasite occurs in the blood. Moranet al. (1999b) showed that direct per osexposure of Atlantic salmon with heavilyinfected tissue did not cause infections. Aswith other myxosporeans, an annelid alter-native host is probably involved in the lifecycle.
Analysis using small subunit ribosomalrDNA (SSU rDNA) suggests that Kudoaspecies are phylogenetically different fromother myxosporean genera examined (i.e.Myxobolus, Henneguya and Myxidium), andthat K. thyrsites in Atlantic salmon is indis-tinguishable from that infecting tubesnout(Aulorhynchus flavidus) and probably othermarine fishes in the Pacific Northwest(Hervio et al., 1997).
Clinical signs and gross pathology. Heavilyinfected fish held on ice for 3–6 days maydevelop extreme softening of the fleshtexture. Occasionally discrete white patchesare visible with the naked eye. The softflesh also will occur following smoking atcool temperatures (below 70°C) where whitepatches in the muscle are readily seen.
Diagnosis. Diagnosis is based on the obser-vation of the characteristic stellate sporesof the parasite, which are about 13 µm indiameter (Fig. 3.10). The spores are bestdetected by microscopic examination of
78 M.L. Kent and T.T. Poppe
Fig. 3.10. Wet mount of Kudoa thyrsites spores. Bar, 15 µm.
88Z:\Customer\CABI\A4337 - Woo\A4419 - Woo Vouchers.vpTuesday, November 05, 2002 11:34:14 AM
Color profile: DisabledComposite Default screen

fluid collected from the freshly cut surfaceof a fillet or by crushing a small pieceof muscle. The parasite shows up wellin Giemsa-stained histological sections.Detection of the parasite in whole fish is aproblem that is, as yet, unresolved.
Wet-mount examination of the hyoidmuscle in the underside of the operculum isa relatively sensitive and specific method fordetecting the infection without damagingthe body musculature (St-Hilaire et al.,1997b). Although this method may missa few light infections, this is not a greatconcern because light infections do notcause soft flesh.
Many copies of the rDNA sequenceoccur within an individual eukaryoticcell, and thus this sequence is useful fordeveloping very sensitive PCR-based tests.Hervio et al. (1997) developed a sensitivePCR test for K. thyrsites, which will be usedto identify the source of infection for salmon.
Control and treatment. There are no com-mercially available drugs against myxo-sporeans. Fish become infected in seawater,so it would be very difficult to eliminateexposure to infections. Because sexuallymature fish and reconditioned grilse aremore prone to the infection, removingsuch fish from the population beforeharvest (e.g. by thorough screening forgrilse in the winter) will greatly minimizethe problem.
Loma salmonae (Microsporidia)
L. salmonae is an obligate intracellularmicrosporidian and infects the gills andother vascularized tissues of salmonidsreared in fresh water (Putz et al., 1965;Putz and McLaughlin, 1970; Morrison andSprague, 1981, 1983; Hauck, 1984; Poynton,1986; Markey et al., 1994; Bruno et al.,1995). Severe gill infections have beenreported in rainbow trout, steelhead troutand kokanee salmon (Wales and Wolf,1955), and Hauck (1984) observed highmortality in chinook due to systemic
infections by a Loma sp. (presumably L.salmonae).
Infections can persist after fish aretransferred to seawater, and the associatedlesions in the gills can become severe in thepen-reared salmon (Kent et al., 1989; Speareet al., 1989). Although the gills are theprimary site of infection, parasites andassociated lesions can occur in the heart,spleen, kidney and pseudobranch. Infectioncan originate in fresh water, but there is ahigh prevalence of L. salmonae in chinooksalmon from netpens that have been rearedsolely on ground water during the fresh-water phase (Kent et al., 1995). Furthermore,infection has been reported in ocean-caughtadult salmon (Kent et al., 1998). Potentialvertical transmission of microsporeans viathe ova has been reported by Vaney andConte (1901) and Summerfelt and Warner(1970). Vertical transmission of L. salmonaefrom infected females to progeny should alsobe considered as a possibility. L. salmonaeoccurs in the ovaries, but not in the eggs,of sexually mature salmon (Docker et al.,1997a). Therefore, the progeny of infectedfemales could become exposed to the para-site through contaminated ovarian fluid.Furthermore, spores of L. salmonae cansurvive iodine treatment at 100 ppm for15 min (Shaw et al., 1999), a dose typicallyused for disinfecting salmonid eggs afterspawning.
Although other Loma species infectnon-salmonid fish, it is doubtful whetherthese fish are reservoirs for L. salmonae.Shaw et al. (1997) demonstrated that aLoma sp. from shiner perch, Cytmatogasteraggregata (a common fish found around seacages in British Columbia), was a differentspecies from L. salmonae. Although thetwo microsporeans were morphologicallyindistinguishable, Shaw et al. (1997)showed that the parasite from shiner perchcould not infect salmon.
Loma morhua is a common gill parasiteof Atlantic cod (Morrison, 1983), and causessimilar gill lesions to those induced by L.salmonae in salmon. Therefore, this parasitealso has potential to cause disease in farmedcod.
Infectious Diseases of Coldwater Fish in Salt Water 79
89Z:\Customer\CABI\A4337 - Woo\A4419 - Woo Vouchers.vpTuesday, November 05, 2002 11:34:14 AM
Color profile: DisabledComposite Default screen

Clinical signs and gross pathology. F i s hwith heavy gill infections are usuallylethargic, and small white cysts may be seenin the gills (Fig. 3.11). A consistent findingin pen-reared salmon is the presence ofmultiple petechiae in an otherwise palegill. Infected gills may also appear nodular.Systemic infections in chinook may causeenlargement of the spleen and kidney. InAtlantic cod, obvious white cysts are alsoapparent in the gills and visceral organs(Morrison, 1983).
Diagnosis. Spores of Loma spp. can beeasily detected in wet-mount preparationsof moderately to heavily infected gills. The
parasite–host cell complexes, xenomas,appear more opaque than the surroundingtissue, and high magnification revealsmasses of the spores within the xenoma(Fig. 3.12). Individual xenomas may occludeblood vessels, and rupture results in amarked inflammatory response by thehost as spores are released (Kent et al., 1989).The spores are bean-shaped and are about5 × 3 µm. Spores in infected tissue areGram-positive.
A sensitive PCR test for L. salmonaeusing an rDNA sequence has been described(Docker et al., 1997a). This may be usefulfor screening fish (i.e. broodstock) forsubclinical infections. Furthermore, these
80 M.L. Kent and T.T. Poppe
Fig. 3.11. Salmon gill with Loma salmonae infection. Note the mottled appearance of the gills andnumerous xenomas (arrowheads).
Fig. 3.12. Wet mount of a gill infected with Loma showing a ruptured xenoma releasing spores(arrowheads). Bar, 10 µm.
90Z:\Customer\CABI\A4337 - Woo\A4419 - Woo Vouchers.vpTuesday, November 05, 2002 11:34:15 AM
Color profile: DisabledComposite Default screen

specific primers can differentiate L.salmonae from other Loma species.
Control and treatment. Currently, there areno licensed pharmacological agents orvaccines (Speare et al., 1998). In laboratorystudies, feeding fumagillin at 10 mg kg−1 fishday−1 for 30 days (Kent and Dawe, 1994)prevented infections in chinook salmon.Our recent experiments demonstrated thatinfections can be controlled with lowerdoses of fumagillin, i.e. 2 or 4 mg kg−1 fish(Kent and Poppe, 1998).
The synthetic analogue of fumagillin,TNP-470 (Takeda Chemical Industries, Ltd,Japan), can also be effective in reducingL. salmonae infections (Higgins et al.,1998). Oral treatment with this compound at0.1 or 1.0 mg kg−1 fish for 4 weeks greatlyreduced the intensity of infections, with noapparent clinical toxic side effects. Speareet al. (1998) showed that rainbow trouthave strong protection against reinfection,which suggests that L. salmonae infectionsmay be prevented by vaccines.
The susceptibility of Pacific salmonstrains to L. salmonae infection usingfeeding trails was examined by Shaw et al.(2000). Differences in strain susceptibilitywere noted and this may assist with futurebreeding experiments.
Nucleospora salmonis
N. salmonis is an unusual microsporidiumthat infects the nuclei of haemoblasts, par-ticularly lymphoblasts or plasmablasts, insalmonid fishes (Chilmonczyk et al., 1991).This microsporidium was first observed inpen-reared chinook in Washington State,where it was associated with anaemia(Elston et al., 1987). The parasite has alsobeen reported in freshwater-reared chinook,kokanee (Oncorhynchus nerka) and steel-head trout (O. mykiss) (Hedrick et al., 1990,1991b; Morrison et al., 1990) The infectionis common in caged-reared chinook salmonin British Columbia and in Atlantic salmonin Chile (Bravo, 1996).
This microsporidium was originallydescribed as N. salmonis (cf. Hedrick et al.,1991a), but was described shortly there-after as Enterocytozoon salmonis byChilmonczyk et al. (1991). Rules of zoo-logical nomenclature, morphological dataand rDNA sequence data support thevalidity of the genus Nucleospora, and itsplacement in the family Enterocytozooidae(see Desportes-Livae et al., 1996; Dockeret al., 1997b). Similar intranuclear micro-sporidia were reported in Atlantic lumpfish(Cyclopterus lumpus) (Mullins et al., 1994)and Atlantic halibut (Nilsen et al., 1995).
N. salmonis infections are usually asso-ciated with a concurrent neoplastic condi-tion involving massive lymphoproliferation,known as plasmacytoid leukaemia (PL) inchinook salmon in British Columbia (Kentet al., 1990). However, the actual cause ofPL is controversial. Laboratory transmissionstudies indicated that N. salmonis maynot be the primary cause of all cases ofPL (Kent and Dawe, 1990; Newbound andKent, 1991), and Eaton and Kent (1992)described a retrovirus associated with thecondition.
It is possible that PL actually representstwo separate diseases; one caused by thevirus and one caused by the micro-sporidium. Studies with fumagillin andTNP-470 (Hedrick et al., 1991b; Higginset al., 1998) support the microsporidianhypothesis, i.e. treatment with theseanti-microsporidian compounds preventedN. salmonis infections and PL. Moreover,in contrast to the late 1980s and early 1990s,N. salmonis is consistently observed in theproliferating plasmablasts in essentially allcases that have been investigated in recentyears in British Columbia.
N. salmonis is transmitted by co-habitation or feeding infected tissues tofish in fresh water (Baxa-Antonio et al.,1992). These findings have been reportedin our laboratory, but we were unable totransmit the infection by cohabitation inseawater. Circumstantial evidence (e.g. theoccurrence of the parasite in Chile) suggeststhat the parasite may be transmitted viaeggs.
Infectious Diseases of Coldwater Fish in Salt Water 81
91Z:\Customer\CABI\A4337 - Woo\A4419 - Woo Vouchers.vpTuesday, November 05, 2002 11:34:15 AM
Color profile: DisabledComposite Default screen

Clinical signs and gross pathology. Heavi lyinfected fish are anaemic. Fish with PL areoften dark, lethargic and may swim near thesurface. Many of the of fish with PL exhibitsevere bilateral exophthalmos (Fig. 3.13).The exophthalmos is due to massive accu-mulation of white or hyperaemic tissue inthe orbit of the eye. The spleen and kidneyare enlarged when systemic infectionsoccur. Petechiae may occur in the liver,mesenteric fat, pancreas, heart and skeletalmuscle. The lower intestinal wall maybe markedly thickened. Some fish have
ascites consisting of a clear or sero-sanguinous fluid.
Diagnosis. This microsporidium is small(about 2 µm) and is identified by carefulexamination of nuclei of haemoblasts inhistological sections or in Gram-stainedimprints (Fig. 3.14). Following Gram stain,the spores stain Gram-positive. They havea characteristic bean shape, and measureabout 2 × 1 µm. Sensitive and specific PCRtests have been developed for the detectionof N. salmonis based on a rDNA sequence
82 M.L. Kent and T.T. Poppe
Fig. 3.13. Severe exophthalmos in chinook salmon with plasmacytoid leukaemia associated withNucleospora salmonis infection.
Fig. 3.14. Gram-stained kidney imprint of Nucleospora salmonis showing spores (arrowhead) in a remnantof a nucleus. Bar, 10 µm.
92Z:\Customer\CABI\A4337 - Woo\A4419 - Woo Vouchers.vpTuesday, November 05, 2002 11:34:16 AM
Color profile: DisabledComposite Default screen

from the small subunit region (Barloughet al., 1995) or internal transcribed spacerregion (Docker et al., 1997b).
Treatment and control. There is no com-mercially available drug for treating N.salmonis infections. However, Hedrick et al.(1991b) controlled the infection in experi-mentally infected chinook by oral treatmentwith fumagillin at 1.0 mg kg−1 fish day−1 for2 weeks. Higgins et al. (1998) found thatthe fumagillin analogue, TNP-470 (TakedaChemical Industries Ltd), was very effectiveat controlling experimental infections whenfish received an oral treatment at either 0.1or 1.0 mg kg−1 fish day−1 for 4 weeks.
Crustacean Parasites
Sea lice – caligid copepods (family Caligidae)
Sea lice are the most economically impor-tant parasites afflicting salmon in cageculture. ‘Sea lice’ refer to several species ofmarine ectoparasitic copepods of the generaLepeophtheirus and Caligus of the familyCaligidae that infect marine fishes, particu-larly salmonids (Costello, 1993; Johnson,1998). Lepeophtheirus salmonis has a cir-cumpolar distribution and is restricted tosalmonids, except as a result of accidentaltransfer from salmonids (Kabata, 1979). Incontrast, Caligus species that infect salmonhave broad host ranges that include bothnon-salmonid teleost and elasmobranchhosts.
Heavy infections greatly reduce themarket value of the fish and ultimately resultin death. Mortality may occur due to thedevelopment of secondary diseases (e.g.vibriosis, furunculosis) exacerbated by thehigh levels of accompanying stress. In severecases where the epidermis is breached,death may be due to a loss of physiologicalhomeostasis including osmotic stress, anae-mia and hypoproteinaemia (Wootten et al.,1982; Tully et al., 1993). Sea lice may alsofunction as vectors of bacteria and virusessuch as infectious salmon anaemia virus(Nylund et al., 1994).
Sea lice have ten developmental stages:two free-living planktonic nauplius stages,one free-swimming infectious copepodidstage, four attached chalimus stages,two pre-adult stages and one adult stage(Johnson and Albright, 1991; Schram, 1993).The copepodid, chalimus, pre-adult andadult stages all feed on mucus, skin andblood of fish (Kabata, 1970; Brandal et al.,1976).
Clinical signs and gross pathology. Pre-adultand adult parasites actively move on thesurface of fish, and lesions caused by thesestages may be severe and widespread.In contrast, damage by the non-motilecopepodid and chalimus larvae is generallyfocal (Bron et al., 1991; Johnson andAlbright, 1992). Infected salmon commonlyhave grey patches (extensive areas of skinerosion) and haemorrhaging on the headand back. They often exhibit distinct areas oferosion, dark coloration and subepidermalhaemorrhage in the perianal region(Wootten et al., 1982; Urawa and Kato, 1991;Nagasawa and Sakamoto, 1993; Johnsonet al., 1996). Severely infected salmon haveulcers in which the epidermis is breachedand the underlying tissues exposed. Theselesions often occur on the head and behindthe dorsal fin (Jónsdóttir et al., 1992).
Diagnosis. Copepodids and chalimuslarvae of sea lice are small (< 4 mm in length)and can occur on the body surface and finsas well as in the buccal cavity and on thegills. Their small size requires the use of amagnifying glass or dissecting microscope todetect their presence.
Pre-adult and adult sea lice are visible tothe naked eye. They are on the body surface,especially on the head, back and in theperianal region. It is these stages that usuallycause the most damage to the fish. Pre-adultand adult stages of Caligus species can bedistinguished from Lepeophtheirus speciesby the presence of lunules on their anteriormargin (Fig. 3.15). There is a key to aidin the identification of adult sea lice ofthe northern hemisphere (Johnson andMargolis, 1994).
Infectious Diseases of Coldwater Fish in Salt Water 83
93Z:\Customer\CABI\A4337 - Woo\A4419 - Woo Vouchers.vpTuesday, November 05, 2002 11:34:17 AM
Color profile: DisabledComposite Default screen

Control and treatment. There are excellentreviews on the control and treatment of sealice (Costello, 1993; Johnson et al., 1993;Roth et al., 1993a; Johnson, 1998). Manage-ment strategies are useful for reducing theimpact of sea lice in cage farms. Farmsshould be located in areas with strong watercurrents to flush away copepodid stages andin areas where wild fish reservoir hosts arenot numerous. Fallowing of sites betweenproduction cycles and maintaining onlysingle year classes at sites can also signifi-cantly reduce the need for treatments forL. salmonis (Bron et al., 1993; Grant andTreasurer, 1993). This approach may beapplied to several sites within a commonarea. In cases where farms belonging todifferent companies are in close proximity toeach other, cooperative agreements betweencompanies with respect to single-year classstocking, periods of fallowing and timingof sea lice treatments have been effectivefor controlling sea lice outbreaks (Grant andTreasurer, 1993).
Chemotherapy has played a significantrole in attempts at controlling sea lice. Bathtreatments with dichlorvos, trichlorfon,azamethiphos, cypermethrin, carbaryl,pyrethroids and hydrogen peroxide havebeen employed (Brandal and Egidius, 1979;
Costello, 1993; Johnson et al., 1993; Rothet al., 1993a,b; Thomassen, 1993a,b). Thesetreatments have mainly been developed fortreating Atlantic salmon, and caution shouldbe used when applying them to otherspecies, as they often vary in their ability totolerate sea lice treatments (see Johnson andMargolis, 1993; Johnson et al., 1993).
The organophosphorus insecticides,dichlorvos, marketed as ‘Nuvan 500EC’or ‘Aquaguard SLT’, or in its relatedtrichlorphon form as ‘Neguvon’, were thefirst chemicals widely used to control sealice (Brandal and Egidius, 1977; Grave et al.,1991a,b). Dichlorvos and trichlorphon havebeen used since the 1960s as a bath treatmentfor parasites in pond fish culture (reviewedin Schmahl et al., 1989). These treatmentseffectively remove both the pre-adult andadult stages of sea lice, but not the chalimuslarvae, from salmon (Brandal and Egidius,1979). Therefore, successive treatments,usually at 2–4 week intervals, are requiredto control infections (Wootten et al., 1982).Repeated treatment with dichlorovos mayinduce resistance in sea lice (Jones et al.,1992).
Another organophosphate, azamethi-phos (marketed as Salmosan® and Alfa-cron®), is presently used in Europe and
84 M.L. Kent and T.T. Poppe
Fig. 3.15. Caligus (a) and Lepeophtheirus (b). Note lunules (arrow) on Caligus. (From Kabata, 1988.)
94Z:\Customer\CABI\A4337 - Woo\A4419 - Woo Vouchers.vpTuesday, November 05, 2002 12:32:03 PM
Color profile: DisabledComposite Default screen

Canada for sea lice control. It is administeredas a bath treatment, and like the otherorganophosphates shows little efficacyagainst the attached chalimus stages (Rothet al., 1996). This chemical is efficaciousagainst L. salmonis, has a wider therapeuticmargin and appears to be more toleratedthan the other organophosphates by Atlanticsalmon (Hodneland et al., 1993; Roth et al.,1993a, 1996).
A drawback with these organophos-phates is that they may be toxic to a widevariety of marine organisms when they arereleased into the surrounding waters aftertreatment. However, their impact on non-target species may be minimal due to dilu-tion and the rapid breakdown of these pesti-cides (Egidius and Moster, 1987; Cusack andJohnson, 1990; Dobson and Tack, 1991).
Pyrethrin and pyrethroid compoundsare currently being used for sea lice control(Boxaspen and Holm, 1991a,b; Roth et al.,1993b). The synthetic pyrethroid cyper-methrin is believed to be more efficaciousthan azamethiphos for the control of L.salmonis on Atlantic salmon. Clinical fieldtrials are ongoing in Maine, USA, usingcypermethrin (under the market nameExcis®). Presently this is the most widelyused bath treatment against sea lice inNorway.
Bath treatments with hydrogenperoxide may also be another effectivemethod for treating sea lice (Bruno, 1992;Thomassen, 1993a,b; Bruno and Raynard,1994). Thomassen (1993a,b) reported thatbath treatments of hydrogen peroxide at aconcentration 1.5 g l−1 for 20 min effectivelyremoves from 85 to 100% of the pre-adultand adult stages of sea lice without beingtoxic to Atlantic salmon. The market name ofhydrogen peroxide for use in fish farming isSalartect 500 FLT®. Twenty minute bathtreatments of 1.5 g l−1 hydrogen peroxide at11°C effectively removed approximately80% of the pre-adult and adult stages ofL. salmonis, but had no significant effect onthe intensity of infection with the attachedchalimus stages. In addition, a high pro-portion of the pre-adult and adult stagesremoved from the fish recovered aftertreatment (Johnson et al., 1993; Bruno and
Raynard, 1994). With respect to L. salmonis,these stages are less likely to reinfect thetreated hosts. Pre-adults and adults ofspecies of Caligus are generally moreactive swimmers and reinfection is possibleif they recover. The use of hydrogen per-oxide is rather impractical and it has beenessentially abandoned in Norway. Further-more, caution should be used when apply-ing hydrogen peroxide treatments, particu-larly at higher temperatures (Johnson et al.,1993; Roth et al., 1993a; Bruno and Raynard,1994). Fortunately, Atlantic salmon are lesssensitive to hydrogen peroxide than Pacificsalmon species, such as chinook salmon(Johnson et al., 1993).
Application of external treatments innetpens is often expensive and difficult.Therefore, considerable effort has beendirected toward development of oraltreatments for sea lice. Palmer et al. (1987)reported the results of preliminary studieson the efficacy of oral doses of ivermectin forthe control of sea lice on Atlantic salmon.Ivermectin has been demonstrated to beeffective in controlling all developmentalstages of sea lice (Smith et al., 1993; Johnsonand Margolis, 1993). Although this drug wasfound to be effective in reducing popu-lations of sea lice, it had a relativelynarrow margin of safety with salmon.Johnson et al. (1993) showed that ivermectincan be very toxic to Atlantic salmon, and thatthe level of toxicity varied between salmonspecies. Atlantic salmon fed 0.05 mg kg−1
on alternate days became anorexic after20 days, and ivermectin was lethal to fishfed at higher doses (Johnson et al., 1993).Fish suffering from ivermectin toxicity arelistless, show ataxia and then die in a fewdays. Due to long tissue withdrawal timesand concerns about the impact of ivermectinresidues in the sediments beneath thenetpens, this drug may never be licensed orregistered for use in aquaculture (Burridgeand Haya, 1993; Costello, 1993).
Oral administration of diflubenzuron(a chemical that inhibits chitin synthesis)reduces infections by both adult and larvalstages of sea lice (Roth et al., 1993a).Treatments using Lepsidon (containingdiflubenzuron) and Ektobann (containing
Infectious Diseases of Coldwater Fish in Salt Water 85
95Z:\Customer\CABI\A4337 - Woo\A4419 - Woo Vouchers.vpTuesday, November 05, 2002 11:34:18 AM
Color profile: DisabledComposite Default screen

teflubenzuron) are being conducted on alimited scale in Norway and the FaroeIslands under special permit. Thesecompounds are highly effective againstthe copepodid, chalimus and pre-adultstages. However, they have no efficacyagainst the adult stages because they nolonger moult. A concern with these drugs isthe possible detrimental effects on non-target arthropods that dwell around seacages. Licensing of these compounds insome areas, such as the USA, may be verydifficult due to laws that limit the use ofdiflubenzuron within 5 km of the coast(Roth et al., 1993a).
Cleaner-fishes (i.e. wrasse species in thefamily Labridae) are used in Norway, theShetland Isles, Scotland and Ireland to con-trol sea lice (reviewed in Costello, 1993;Kvenseth, 1993; Treasurer, 1993; Tully et al.,1996). In laboratory and field studies, wrasseremove sea lice from salmonids but notalways in a predictable manner. A surveyof fish farmers in Scotland who have triedwrasses to control sea lice showed that themajority felt that their use was beneficial,particularly when used in conjunctionwith dichlorvos treatments (Anon., 1991).Wrasses are also useful for reducing foulingon cages. The disadvantages of using wrasseto control sea lice include the requirementfor smaller mesh size in nets to prevent theirescape, intimidation by larger salmon anda tendency not to clean them, aggressivebehaviour and infliction of scale and eyedamage to the salmon (Anon., 1991). Inaddition, wrasse exhibit high over-wintermortalities, their supply is limited and theircost is usually high. Nevertheless, wrasseare used in over half of the fish farms inNorway.
A vaccine for sea lice would be useful.Although Atlantic salmon can produce anti-bodies to sea lice extracts in controlledstudies (Grayson et al., 1991), salmon natu-rally infected with L. salmonis and Caliguselongatus fail to produce an antibodyresponse (Grayson et al., 1991; MacKinnon,1991). Furthermore, there was no differencein the number of copepods carried oncontrol and immunized Atlantic salmon
when exposed under laboratory conditions(Grayson et al., 1995).
Family Pennellidae
Members of the family Pennellidae maycause problems in netpen aquaculture.Haemobaphes disphaerocephalus has beenreported in pen-reared Atlantic salmon(Kent et al., 1997). This parasitic copepodnormally infects eulachon (Thaleichthyspacificus) and this was the first report ofa Haemobaphes species infecting salmon.The parasite penetrates the branchial vascu-lature and causes anaemia. Fortunately, theinfection has been observed in only a fewAtlantic salmon reared in British Columbia.In contrast, Lernaeocera branchialis is acommon parasite of Atlantic cod (Kabata,1984). The parasite causes reduced growthand anaemia (Khan, 1988), and Khan et al.(1990) concluded that it is a potential threatto cod farming.
Clinical disease and gross pathology. Fishinfected with either Haemobaphes or Lern-aeocera are usually anaemic and may belethargic. Examination of the opercula cav-ity reveals the coiled egg sacs and blood-engorged body of the parasite penetratingthe gill (Fig. 3.16). The long neck and ante-rior holdfast are internal within the gill arch.
Diagnosis. Haemobaphes and Lernaeoceraspp. are characterized by attachment at thegill arch and coiled egg sacs. Specific identi-fication requires examination of the anteriorholdfast (see Kabata, 1988), which must bevery carefully dissected from tissues. Atlan-tic lumpfish (C. lumpus) is an intermediatehost of L. branchialis, and thus simultaneousrearing of this fish with cod should bediscouraged.
Control and treatment. There is no suitabledrug available for treating this infection. Theinfective larvae are free-swimming and itwould be difficult to prevent the infection innetpens.
86 M.L. Kent and T.T. Poppe
96Z:\Customer\CABI\A4337 - Woo\A4419 - Woo Vouchers.vpTuesday, November 05, 2002 11:34:18 AM
Color profile: DisabledComposite Default screen

Isopods – Ceratothoa gaudichaudii
Four species of isopods, Ceratothoagaudichaudii, Rocinela maculata, Rocinelabelliceps pugettensis and Gnathia sp.,have been reported from seawater-rearedsalmonids (Novotny and Mahnken, 1971;Awakura, 1980, 1983; Drinan and Rodger,1990; Inostroza et al., 1993). The onlyeconomically important isopod parasite ofmarine coldwater netpen-reared fish hasbeen C. gauchichaudii. In Chile, C.gaudichaudii has been reported from awide variety of native hosts. This lowhost specificity has allowed this parasiteto successfully infect coho and Atlanticsalmon. Disease caused by this parasite hasbeen a serious problem at certain farm sitesin Chile (Inostroza et al., 1993).
Clinical signs and gross pathology. C. gaudi-chaudii feeds on host blood, attaching tothe inner mouth surfaces and less frequentlyto the gills (Sievers et al., 1995). Diseaseis caused by their attachment and feedingactivities. Damage to the host includessevere erosion of gill lamellae and ulcers onthe gill arch and inside the mouth.
Diagnosis. Ceratothoa is readily identifiedon the fish by examination of the mouth andgills (Fig. 3.17).
Control and treatment. Sievers et al. (1995)evaluated the efficacy of eight commercialinsecticides against C. gaudichaudii on
Infectious Diseases of Coldwater Fish in Salt Water 87
Fig. 3.16. Haemobaphes disphaerocephalus infecting the gills of an Atlantic salmon.
Fig. 3.17. Ceratothoa gaudichaudii on the gills ofAtlantic salmon (courtesy of R. Inostroza).
Atlantic salmon. Sixty-minute bathtreatments with the organophosphates,trichlorfon (Neguvon) and dichlorvos(Nuvan 1000) at concentrations of 300and 3 ppm, respectively, were found to be100% effective against this parasite withouttoxicity to the fish.
Helminth Parasites
Fish are infected with a wide variety ofparasitic worms, collectively referred toas helminth parasites. Although theseparasites are very common in wild fish,and occasionally infect cultured species,generally they do not cause severe disease.However, certain helminths can causedamage when infections are heavy or whenthey infect a critical organ. In addition,
97Z:\Customer\CABI\A4337 - Woo\A4419 - Woo Vouchers.vpTuesday, November 05, 2002 11:34:19 AM
Color profile: DisabledComposite Default screen

some helminth parasites of fish can infecthumans or cause unsightly lesions. Thefollowing are helminth parasites of impor-tance in coldwater marine netpen-rearedfish.
Cestodes (tapeworms)
Two life stages of cestodes are found infish: adults infect the digestive tract andmetacestodes (juveniles) are usually on theinternal organs or muscle. The first interme-diate hosts of tapeworms that infect fish areusually crustaceans (e.g. copepods). Fishmay also be the second intermediate hostsfor tapeworms, and a fish-eating mammal orbird, or another fish, are definitive hosts.Therefore, fish usually acquire metacestodeinfections by eating infected crustaceans.Metacestodes in fish tissues often cause aninflammatory response to the encapsulatedor migrating parasite. The only reportedsignificant metacestode disease of salmonreared in marine netpens is caused byGilquinia squali, which infects the eyes ofchinook salmon (Kent et al., 1991).
Adult cestodes – Eubothrium spp.
Eubothrium spp. are common cestode para-sites of salmonid fish in both fresh and saltwater, in which adults develop in the gut.Infections with one species in pen-reared
Atlantic salmon in Norway have beenassociated with reduced growth and, occa-sionally, mortality (Bristow and Berland,1991a,b; Håstein and Lindstad, 1991). Theweight of infected market-size farmedAtlantic salmon in Norway is 10–15% lessthan uninfected salmon (Berland andBristow, 1994). It has been noted that asimilar infection occurs in the cestodesof broodstock from pen-reared chinooksalmon in British Columbia.
The fish acquire infections ofEubothrium species by ingesting first inter-mediate hosts (presumably copepods)infected with the procercoid stage, orpossibly transport hosts infected withplerocercoids. The life cycle of thistapeworm in marine fishes has not beenelucidated, but its freshwater counterpart,Eubothrium salvelini, uses copepods(Cyclops spp.) as its intermediate host.Procercoids that develop in Cyclops aredirectly infective for juvenile salmon(Boyce, 1974). E. salvelini is known to affectsurvival, growth and stamina, and to haveother debilitating effects on juvenile sockeyesalmon (Boyce and Behrens-Yamada, 1977;Boyce, 1979; Boyce and Clarke, 1983).
Clinical signs and gross pathology. Heavi lyinfected fish are often smaller than aver-age. Dissection of the gut will reveal num-erous, white, flat, ‘tape-like’ worms in theintestine and pyloric caeca (Fig. 3.18). Heavy
88 M.L. Kent and T.T. Poppe
Fig. 3.18. Adult Eubothrium cestodes in the gut of Atlantic salmon (courtesy of B. Berland).
98Z:\Customer\CABI\A4337 - Woo\A4419 - Woo Vouchers.vpTuesday, November 05, 2002 11:34:19 AM
Color profile: DisabledComposite Default screen

infections may induce anaemia, and whenextremely severe the cestodes may causedeath due to blockage of the intestinal tract(Mitchell, 1993).
Diagnosis. Adult cestodes are usually long,flat, whitish and segmented. Identificationas a cestode is based on a segmented body, ascolex (anterior end), and on the structureand arrangement of the reproductive systemwithin the segments (Schmidt, 1986; Khalilet al., 1994). Eubothrium lacks hooks on thescolex, which is elongate with two shallowgrooves – one dorsal and the other ventral.
Control and treatment. Oral treatment foradult tapeworms with anthelmintic drugs,such as praziquantel, may be effective(Mitchell, 1993). Avoiding cestode infec-tions is difficult because infected inter-mediate hosts (i.e. crustaceans) move freelythroughout netpens.
Gilquinia squali metacestodes
Eye infections by metacestodes of G. squali(order Trypanorhyncha) have been associ-ated with mortality of young chinooksalmon at netpen sites in British Columbia(Kent et al., 1991). The definitive host forG. squali is the spiny dogfish (Squalusacanthus). The metacestode is common inthe eyes of North Sea whiting, Merlangiusmerlangus (MacKenzie, 1965, 1975). Wildsalmon are probably not a normal inter-mediate host for the worm as this infectionhas only been seen in cage-reared salmon(Kent et al., 1991).
The adult worms are prevalent indogfish throughout marine waters in BritishColumbia, and gravid adult worms arecommonly found in dogfish in the spring.Dogfish are frequently found in or aroundnetpens at this time, thus providing infectivecoracidia for the first intermediate host nearthe pens. Based on trypanorhynchs whoselife cycles have been investigated (Mundryand Dailey, 1971; Overstreet, 1978; Mattis,1986; Sakanari and Moser, 1989), it is likelythat a crustacean is the first intermediatehost for G. squali. Chinook salmon in
netpens presumably acquire the infection byeating this crustacean.
Clinical signs and gross pathology. M o r i-bund fish are lethargic and remain near thebottom of the pen. The fish can easily becaptured by hand, suggesting that they areblind. However, fish are not usually emaci-ated and contain food in their stomachs,which indicates that they continue to feed.
The lens often appears normal, whereasthe iris and vitreous chamber may be whiteand opaque and occasionally haemorrhagic.The lens in more severely affected eyes isalso opaque, suggesting cataracts. In extremecases, the globe is ruptured and the lens isextruded. However, many fish die with theglobe of the eye still intact. The eye lesionsare often bilateral.
Diagnosis. The infection is identified bydetecting trypanorhynch metacestodes inthe vitreous humour. Trypanorhynch ces-todes are identified by the presence of fourreversible, spiny tentacles, which emergefrom the apex of the scolex (Fig. 3.19).
Control and treatment. There is no knowntreatment for Gilquinia infection in fish.Infestation can only be controlled by pre-venting infection. Fish that are feeding wellon commercial diets and thus feed less onnatural biota appear to have lower infec-tions. The complete life cycle of the parasiteis unknown, so precise recommendationsfor avoiding infection are not available.Furthermore, preventing transmission ofthe parasite from dogfish to salmon viathe arthropod first intermediate hosts wouldbe difficult because of uncontrolled watermovement into netpens and unrestrictedmovement of dogfish around netpens.
Digenetic trematodes (flukes)
As with cestodes, fish can be intermediatehosts or definitive hosts for digenetictrematodes. Almost all flukes have eithera two-host or three-host life cycle, butthere is a wide variety of life cycle patterns
Infectious Diseases of Coldwater Fish in Salt Water 89
99Z:\Customer\CABI\A4337 - Woo\A4419 - Woo Vouchers.vpTuesday, November 05, 2002 11:34:20 AM
Color profile: DisabledComposite Default screen

(Williams and Jones, 1994). With a fewexceptions among the marine fish bloodflukes of the family Sanguinicolidae,molluscs (either snails or bivalves) are thefirst intermediate hosts of digeneans andsecond intermediate hosts may be an inver-tebrate or a fish. Except in occasionalcircumstances where the life cycle has beenforeshortened, the definitive hosts are verte-brates. The species of concern in marinefarming of salmon use either birds or fish asdefinitive hosts, with the fish serving as thesecond intermediate host. Cercariae of thesespecies emerge from molluscs and infectthe salmonid host by direct penetration ofthe skin or gills, subsequently developinginto a resting stage known as a metacercaria,which may be encysted or unencysteddepending upon the final site of infection inthe fish.
Heavy infections by metacercariae are ofconcern because they can cause morbidity.In addition, metacercarial infections of theskin or muscle can be important becausethey may reduce the aesthetic quality of thefish. Except for blood flukes and a group oftissue parasites of the family Didymozoidaefound mainly in scombroid fishes, mostadult flukes of fish infect the alimentary tractand seldom cause significant tissue damage.
The metacercariae of four digeneantrematodes have caused problems inseawater pen-reared salmonid fishes:
‘neascus’-type, Diplostomum sp., Crypto-cotyle lingua and Stephanostomum tenue.
Skin diseases caused by digenean trematodes
Black grub (larval type neascus) is themetacercarial stage of certain species ofthe family Diplostomatidae, which includesseveral genera (Gibson, 1996). This parasiteinfects a wide variety of freshwater fishes,including salmonids. Freshwater snails arethe first intermediate hosts, and cercariaereleased from infected snails penetratebeneath the scales in the dermis of the fishhost. Fish-eating birds serve as the defini-tive hosts. These infections can persist afterfish are transferred to seawater. The prob-lem has been recorded in pen-reared cohosalmon in British Columbia. A conditionsimilar to black grub in coho salmon is‘black spot disease’ in Atlantic salmoncaused by metacercariae of C. lingua. Incontrast to the former condition, this para-site has a marine life cycle, which involvesa definitive (adult) stage in fish-eatingbirds and a cercarial stage in snails. ACryptocotyle sp. has also caused similarinfections in cage-reared Atlantic cod(Lysne et al., 1994).
Clinical signs and gross pathology. Infectedfish exhibit few to numerous raised blackspots up to 1 mm in diameter in the skin,
90 M.L. Kent and T.T. Poppe
Fig. 3.19. Gilquinia squali metacestode from the eye of a chinook salmon. Semichon’s acetocarmine. Bar,0.2 mm.
100Z:\Customer\CABI\A4337 - Woo\A4419 - Woo Vouchers.vpTuesday, November 05, 2002 11:34:20 AM
Color profile: DisabledComposite Default screen

fins, cornea and gills (Fig. 3.20). Occa-sionally, encysted metacercariae may befound in internal organs. Fish seldombecome clinically affected unless they areheavily infected.
Diagnosis. Diagnosis of metacercarialinfections in general is relatively easy usingwet mounts or histological sections. How-ever, more precise identification to thegenus or species level usually requirescareful preparations of the metacercariae instained whole mounts and examination ofthe internal anatomy (Fig. 3.21). Informationon the first occurrence of the infection(i.e. marine vs. freshwater) is useful fordifferentiating neascus from Cryptocotyle.
Control and treatment. As the cercarialstage of Cryptocotyle is common in theperiwinkle (Littorina littorea), cages locatedin shallow water and close to the shoreare more prone to the infection. There isno known treatment for this infection.With neascus, fish become infected infresh water by exposure to surface watercontaining infected snails. Disinfection ofthe water supply or using ground watershould eliminate or greatly reduce theinfection. Based on reports from one fishfarm in British Columbia, removing fishfrom freshwater hatcheries and introducingthem to netpens before June or July mayreduce the intensity and prevalence of theinfection.
Stephanostomum heart infections
Heart (pericardial cavity) infections bymetacercariae of S. tenue caused high
Infectious Diseases of Coldwater Fish in Salt Water 91
Fig. 3.20. Cryptocotyle (black spot) metacercariae in the skin of a Atlantic salmon.
Fig. 3.21. Cryptocotyle metacercaria. (FromGibson, 1996.)
101Z:\Customer\CABI\A4337 - Woo\A4419 - Woo Vouchers.vpTuesday, November 12, 2002 3:09:30 PM
Color profile: DisabledComposite Default screen

mortalities in pen-reared rainbow trout inAtlantic Canada (McGladdery et al., 1990).Rainbow trout are accidental hosts for thismarine fluke, which normally infectsmummichog (Fundulus heteroclitus) orsilverside (Menidia menidia) as secondintermediate hosts. Teleost fish, such asthe American eel (Anguilla rostrata), arethe definitive host for the parasite in thevicinity of the affected netpen sites. Thefirst intermediate host is the mud dog whelk(Nassarius obsoletus), which is commonaround the affected netpens.
Clinical signs and gross pathology. T h einfection was associated with high mor-talities in the summer months when watertemperatures increase, presumably due tocardiac dysfunction (McGladdery et al.,1990).
Control and treatment. M a i n t a i n i n gnetpens in water with over 7 m clearancefrom the bottom may reduce the intensity ofinfection (McGladdery et al., 1990).
Nematodes
Nematodes (roundworms) are commonparasites of fish, and occasionally infectpen-reared salmon. As with cestodes anddigenetic trematodes, fish can be eitherdefinitive or intermediate hosts for nema-todes. Crustaceans and, less frequently,other invertebrates are the first intermediatehosts for nematodes that infect fish.
Members of the family Anisakidae arethe only important (and reported) nema-todes of pen-reared salmon. The nematodethat has been associated with disease insalmon netpens is Hysterothylaciumaduncum, which in its adult stage infects thefish digestive tract.
Large numbers of adult Hysterothy-lacium (= Thynnascaris) aduncum werefound blocking the anterior part of the intes-tine of pen-reared rainbow trout in Norwaysome months after the trout were fed freshwild sprat (Sprattus sprattus) that containedjuvenile H. aduncum in their viscera
(Berland, 1987). Intestinal infections ofseapen-reared coho salmon and rainbowtrout with Hysterothylacium spp. havealso been observed in Chile (González andCarvajal, 1994).
Calanoid copepods are the first interme-diate host of H. aduncum, whereas fish andvarious invertebrates, such as polychaetes,barnacles and amphipods, act as secondintermediate hosts (Svendsen, 1990).
Clinical signs and gross pathology. Carvajalet al. (1990) suggested that heavily infectedfish exhibit poor growth, and Berland andEgidius (1980) have attributed mortalitiesin pen-reared rainbow trout in Norwayto heavy intestinal infections with H.aduncum.
Diagnosis. Diagnosis is based on the identi-fication of the worm. Presumptive diagnosisis based on the observation of nematodesin gut lumen. The worms are whitish andcylindrical, and adults are about 40–80 mmin length (Gonzáles and Carvajal, 1994).Confirmation of identification requiresmicroscopical examination of cleared ordissected worms for certain pathognomonicanatomical features of the worm’s digestivetract (Möller and Anders, 1986; Berland,1989). Hysterothylacium and Anisakisspp. can be differentiated from Pseudo-terranova and Contracaecum in that theyhave straight digestive tracts, without aventricular appendix or intestinal caecum.Hysterothylacium can be separated fromAnisakis in that the former has the excretorypore at the level of the nerve ring, whereasit occurs near the anterior tip with Anisakis.
Control and treatment. To prevent infec-tions with Hysterothylacium or Contracae-cum spp., or to keep infection low, the use offresh wild marine fish or fish offal as feed forfarmed fish should be avoided (Berland andEgidius, 1980; Vismanis et al., 1984). Thesefish are intermediate or paratenic (transport)hosts for the nematodes. González andCarvajal (1994) also referred to use of anthel-mintics to reduce infections but did notspecify the anthelmintics they employed.
92 M.L. Kent and T.T. Poppe
102Z:\Customer\CABI\A4337 - Woo\A4419 - Woo Vouchers.vpTuesday, November 05, 2002 11:34:22 AM
Color profile: DisabledComposite Default screen

Acknowledgements
We thank our colleagues with specificexpertise in the following subjects for theiradvice: Dr P. Bustos, ADL Diagnostics ChileLtda, for the P. salmonis information forChile; Dr T.P.T. Evelyn, Canada Fisheriesand Oceans, for general information onbacterial diseases; and Dr S.C. Johnson,Canadian National Research Council, forsea lice information.
References
Almendras, F.E. (1996) Transmission andpathogenesis of Piscirickettsia salmonis inAtlantic salmon (Salmo salar). MSc Thesis,Atlantic Veterinary College, Charlottetown,Prince Edward Island, Canada.
Almendras, F.E. and Carmen Fuentealba, I. (1997)Salmonid rickettsial septicaemia caused byPiscirickettsia salmonis: a review. Diseasesof Aquatic Organisms 29, 137–144.
Amin, A. and Trasti, J. (1988) Endomyocarditisin Atlantic salmon in Norwegian seafarms.A case report. Bulletin of the EuropeanAssociation of Fish Pathologists 8, 70–71.
Anderson, J.I.W. and Conroy, D.A. (1969)The pathogenic myxobacteria with specialreference to fish diseases. Journal of AppliedBacteriology 32, 30–39.
Anderson, J.W. and Conroy, D.A. (1970) Vibriodisease in fishes. In: Snieszko, S.F. (ed.) Dis-eases of Fishes and Shellfishes. AmericanFisheries Society Special Publication 5,Washington, DC, pp. 266–272.
Anon. (1991) Scottish farmers take a long view onlouse control. Fish Farming International 14,45.
Anon. (1996) Proceedings of the MouthrotWorkshop, 12 October 1995, Campbell River,British Columbia. BC Ministry of Agri-culture, Fisheries, and Food, CommericalFisheries Branch, Seafood Industry Develop-ment Report 96-01, 26 pp.
Armstrong, R., Robinson, J., Rymes, C. andNeedham, T. (1993) Infectious haemato-poietic necrosis in Atlantic salmon in BritishColumbia. Canadian Veterinary Journal 34,312–313.
Awakura, T. (1980) On the parasites and parasiticdisease of salmonid fish in Hokkaido. FishPathology 14, 207–209 (in Japanese).
Awakura, T. (1983) Fish disease in Hokkaido – 23.Rocinellosis of coho salmon. Uo to Mizu 21,28–29 (in Japanese).
Barja, J.L. and Toranzo, A.E. (1993) Myolique-faction post-mortem caused by the myxo-sporean Kudoa thyrsites in reared Atlanticsalmon in Spain. Bulletin of the EuropeanAssociation of Fish Pathologists 13, 86–88.
Barlough, J.E., McDowell, T.S., Milani, A.,Bigornia, L., Slemenda, S.B., Pieniazek, N.J.and Hedrick, R.P. (1995) Nested polymerasechain reaction for detection ofEnterocytozoon salmonis genomic DNA inchinook salmon Oncorhynchus tshawytscha.Diseases of Aquatic Organisms 23, 17–23.
Baudin-Laurencin, F. and Bennassr, N. (1993)Post-mortem liquefaction of seawater farmedbrown trout Salmo trutta resulting fromKudoa infections. In: Diseases of Fish andShellfish. Book of Abstracts, Sixth Interna-tional Conference of the European Associa-tion of Fish Pathologists, 5–10 September1993, Brest, France, p. 60.
Baxa, D.V., Kawai, K. and Kusuda, R. (1986) Char-acteristics of gliding bacteria isolated fromdiseased cultured flounder, Paralichthysolivaceus. Fish Pathology 21, 251–258.
Baxa, D.V., Kawai, K. and Kusuda, R. (1987) Exper-imental infection of Flexibacter maritimusin black seabream (Acanthopagrus schlegeli)fry. Fish Pathology 22, 105–109.
Baxa-Antonio, D., Groff, J.M. and Hedrick, R.P.(1992) Experimental horizontal transmissionof Enterocytozoon salmonis to chinooksalmon, Oncorhynchus tshawytscha. Journalof Protozoology 39, 699–702.
Becker, C.D. (1977) Flagellate parasites of fish.In: Kreier, J.P. (ed.) Parasitic Protozoa, Vol. 1Academic Press, New York, pp. 358–416.
Berland, B. (1987) Helminth problems insea-water aquaculture. In: Stenmark, A. andMalmberg, G. (eds) Parasites and Diseasesin Natural Waters and Aquaculture inNordic Countries. Zoo-Tax, NaturhistoriskaRiksmuseet, Stockholm, pp. 56–62.
Berland, B. (1989) Identification of larvalnematodes from fish. In: Moller, H. (ed.)Nematode Problems in North Atlantic Fish.Report from a Workshop in Kiel, 3–4 April1989. International Council for the Explora-tion of the Sea C.M./F:6, pp. 16–22.
Berland, B. and Bristow, G.A. (1994) The cestodeEubothrium sp. in farmed marine salmon. In:Abstracts of the 8th International Congress ofParasitology, Vol. 1, 10–14 October 1994,Izmir, Turkey, p. 53.
Infectious Diseases of Coldwater Fish in Salt Water 93
103Z:\Customer\CABI\A4337 - Woo\A4419 - Woo Vouchers.vpTuesday, November 05, 2002 11:34:22 AM
Color profile: DisabledComposite Default screen

Berland, B. and Egidius, E. (1980) Rundmark ioppdrettsfisk – større problem enn antatt?Norsk Fiskeoppdret. Medlemsblad forNorske Fiskeoppdretteres Forening 5, 16(in Norwegian).
Bernardet, J.F., Campbell, A.C. and Buswell, J.A.(1990) Flexibacter maritimus is the agentof ‘black patch necrosis’ in Dover sole inScotland. Diseases of Aquatic Organisms 8,233–237.
Bernardet, J.-F., Segers, P., Vancanneyt, M.,Berthe, F., Kersters, K. and Vandamme, P.(1996) Cutting a Gordian knot: emendedclassification and description of the genusFlavobacreium, emended description of thefamily Flavobacteriaceae, and proposal ofFlavobacterium hydatis nom. nov. (basonym,Cytophaga aquatilis Strohl and Tait 1978).International Journal of Systematic Bacteri-ology 46, 128–148.
Bloch, B., Gravningen, K. and Larsen, J.L. (1991)Encephalomyelitis among turbot associatedwith a picornavirus-like agent. Diseases ofAquatic Organisms 10, 65–70.
Borg, A. (1960) Studies on myxobacteriaassociated with diseases in salmonidfishes. Wildlife Diseases (microfiche) 8, 1–85(2 microcards).
Boxaspen, K. and Holm, J.C. (1991a) New biocidesused against sea lice compared to organo-phosphorous compounds. In: De Pauw,N. and Joyce, J. (eds) Aquaculture andthe Environment. European AquacultureSociety Special Publication No. 16, Belgium,pp. 393–402.
Boxaspen, K. and Holm, J.C. (1991b)Blomsterekstrakt som avslusingsmiddel forlaks [Extract from chrysanthemum flowersas a delousing agent against salmon lice].Fiskets Gang 77, 17–19.
Boyce, N.P.J. (1974) Biology of Eubothriumsalvelini (Cestoda: Pseudophyllidae), aparasite of juvenile sockeye salmon(Oncorhynchus nerka) of Babine Lake,British Columbia. Journal of the FisheriesResearch Board of Canada 31, 1735–1742.
Boyce, N.P. (1979) Effects of Eubothrium salvelini(Cestoda: Pseudophyllidea) on the growthand vitality of sockeye salmon, Oncor-hynchus nerka. Canadian Journal of Zoology57, 597–602.
Boyce, N.P. and Behrens-Yamada, B. (1977)Effects of a parasite, Eubothrium salvelini(Cestoda: Pseudophyllidea), on the resis-tance of juvenile sockeye salmon, Oncorhyn-chus nerka, to zinc. Journal of the FisheriesResearch Board of Canada 34, 706–709.
Boyce, N.P. and Clarke, W.C. (1983) Eubothriumsalvelini (Cestoda: Pseudophyllidea) impairsseawater adaptation of migrant sockeye year-lings (Oncorhynchus nerka) from BabineLake, British Columbia. Canadian Journal ofFisheries and Aquatic Sciences 40, 821–824.
Brandal, P.O. and Egidius, E. (1977) Preliminaryreport on oral treatments against salmon liceLepeophtheirus salmonis, with Neguvon.Aquaculture 18, 177–178.
Brandal, P.O. and Egidius, E. (1979) Treatmentof salmon lice (Lepeophtheirus salmonisKrøyer, 1838) with Neguvon® – descriptionof method and equipment. Aquaculture 18,183–188.
Brandal, P.O., Egidius, E. and Romslo, I. (1976)Host blood: a major food component forthe parasitic copepod Lepeophtheirussalmonis Krøyer, 1838 (Crustacea:Caligidae). Norwegian Journal of Biology 24,341–343.
Branson, E.J. and Nieto Diaz-Munoz, D. (1991)Description of a new disease condition occur-ring in farmed coho salmon, Oncorhynchuskisutch (Walbaum), in South America.Journal of Fish Diseases 14, 147–156.
Bravo, S. (1994) First report of Piscirickettsiasalmonis in freshwater. Fish Health Section/American Fisheries Society Newsletter 22, 6.
Bravo, S. (1996) Enterocytozoon salmonis inChile. Fish Health Section/AmericanFisheries Society Newsletter 24, 12–13.
Bristow, G.A. and Berland, B. (1991a) A reporton some metazoan parasites of wild marinesalmon (Salmo salar L.) from the west coast ofNorway with comments on their interactionswith farmed salmon. Aquaculture 98,311–318.
Bristow, G.A. and Berland, B. (1991b) The effectof long term, low level Eubothrium sp.(Cestoda: Pseudophyllidea) infection ongrowth of farmed salmon (Salmo salar L.).Aquaculture 98, 325–330.
Brocklebank, J.R., Speare, D.J., Armstrong, R.D.and Evelyn, T.P.T. (1992) Septicemia infarmed Atlantic salmon and chinook salmondue to a rickettsia-like agent. CanadianVeterinary Journal 34, 745–748.
Bron, J.E., Sommerville, C., Jones, M. and Rae,G.H. (1991) The settlement and attachmentof early stages of the salmon louse,Lepeophtheirus salmonis (Copepoda:Caligidae) on the salmon host Salmo salar.Journal of Zoology 224, 201–212.
Bron, J.E., Sommerville, C., Wootten, R. and Rae,G.H. (1993) Fallowing of marine Atlanticsalmon, Salmo salar L., farms as a method for
94 M.L. Kent and T.T. Poppe
104Z:\Customer\CABI\A4337 - Woo\A4419 - Woo Vouchers.vpTuesday, November 05, 2002 11:34:22 AM
Color profile: DisabledComposite Default screen

the control of sea lice, Lepeophtheirussalmonis (Krøyer, 1837). Journal of FishDiseases 16, 487–493.
Bruno, D.W. (1992) The use of hydrogen peroxide(H2O2) to remove sea lice from farmedAtlantic salmon. In: First European Con-ference on Crustacea, September 1992,Paris, p.19.
Bruno, D.W. and Raynard, R.S. (1994) Studies onthe use of hydrogen peroxide as a method forthe control of sea lice on Atlantic salmon.Aquaculture International 2, 10–18.
Bruno, D. and Poppe, T.T. (1996) A Colour Atlasof Salmonid Diseases. Academic Press,London, 194 pp.
Bruno, D.W., Hastings, T.S. and Ellis, A.E. (1986)Histology, bacteriology, and experimental.transmission of cold water vibriosis inAtlantic salmon Salmo salar. Diseases ofAquatic Organisms 1, 163–168.
Bruno, D.W., Collins, R.O. and Morrison, C.M.(1995) The occurrence of Loma salmonae sp.,(Protozoa: Microspora) in farmed rainbowtrout, Oncorhynchus mykiss Walbaum, inScotland. Aquaculture 133, 341–344.
Burridge, L.E. and Haya, K. (1993) The lethality ofivermectin, a potential agent for treatment ofsalmonids against sea lice, to the shrimpCragnon septemspinosa. Aquaculture 117,9–14.
Bustos, P., Entrala, P., Montana, J. andCalbuyahue, J. (1994) Septicemia rickettsialsalmonídea (SRS): éstudio de transmisiónvertical en salmón coho (Oncorhynchuskistuch). Seminario Internacional, Patologíay Nutrición en el desarrollo de laAcuicultura: Factores de éxito, FundaciónChile, 3–7 October 1994, Puerto Montt, Chile.
Cameron, D.E. (1993) Amoebic gill diseasefield research 1992/93. In: Proceedings of theSaltas Research Review Seminar, 29 April1993, Hobart, Australia, pp. 89–102.
Cameron, D.E. (1994) AGD Field Research1993–94. III. Field testing of hydrogen per-oxide with AGD-infected Atlantic salmon:– efficacy and toxicity. In: Report of theSaltas 1993/94 Reseach and DevelopmentProgramme, Hobart, Australia, pp. 119–120.
Carvajal, J., Ruiz, G. and González, L. (1990)Presencia de Hysterothylacium sp.(Nematoda: Anisakidae) en salmón coho deChiloé cultivado en jaulas. Revista Chilenade Historia Natural 63, 165–168.
Chilmonczyk, S., Cox, W.T. and Hedrick, R.P.(1991) Enterocytozoon salmonis n. sp.: anintranuclear microsporidium from salmonidfish. Journal of Protozoology 38, 264–269.
Christie, K.E. (1996) Immunization with viraland parasite antigens: infectious pancreaticnecrosis. In: Book of Abstracts, InternationalSymposium of Fish Vaccinology, 5–7 June1996, Oslo.
Colwell, R.R. and Grimes, D.J. (1984) Vibriodiseases of marine fish populations.Helgolaender Wissenschaftliche Meeresun-tersuchungen 37, 265–287.
Costello, M.J. (1993) Review of methods to controlsea lice (Caligidae: Crustacea) infestations ofsalmon (Salmo salar) farms. In: Boxshall,G.A. and DeFaye, D. (eds) Pathogens of Wildand Farmed Fish: Sea Lice. Ellis Horwood,Chichester, UK, pp. 219–252.
Cusack, R. and Johnson, G. (1990) A studyof dichlorvos (Nuvan, 22,dichloro-ethenyldimethyl phosphate) a therapeuticagent for the treatment of salmonid infectedwith sea lice (Lepeophtheirus salmonis).Aquaculture 90, 101–112.
Cvitanich, J.D., Garate, O. and Smith, C.E. (1991)The isolation of a rickettsia-like organismcausing disease and mortality in Chileansalmonids and its confirmation by Koch’spostulate. Journal of Fish Diseases 14,121–145.
Cvitanich, J.D., Gárate, N.O., Silva, P.C., Andrade,V.M., Figueroa, P.C. and Smith, C.E. (1995)Isolation of a new rickettsia-like organismfrom Atlantic salmon in Chile. FishHealth Section/American Fisheries SocietyNewsletter 23, 1–3.
Dannevig, B.H., Falk, K. and Namork, E. (1995)Isolation of the causal virus of infectioussalmon anaemia (ISA) in a long-term cell linefrom Atlantic salmon head kidney. Journal ofGeneral Virology 76, 1353–1359.
Desportes-Livae, I., Chilmonczyk, S., Hedrick, R.,Ombrouck, C., Monge, D., Maiga, I. andGentilini, M. (1996) Comparative develop-ment of two microsporidian species: Entero-cytozoon bieneusi and Enterocytozoonsalmonis, reported from AIDS patients andsalmonid fish, respectively. Journal ofEukaryotic Microbiology 43, 49–60.
Dobson, D.P. and Tack, T.J. (1991) Evaluationof the dispersal of treatment solutions ofdichlorvos from marine salmon pens.Aquaculture 95, 15–32.
Docker, M.F., Devlin, R.H., Richard, J. and Kent,M.L. (1997a) Sensitive and specific polymer-ase chain reaction assay for detection of Lomasalmonae (Microsporea). Diseases of AquaticOrganisms 29, 13–20.
Docker, M.F., Kent, M.L., Hervio, D.M.L., Weiss,L., Cali, A. and Devlin, R.H. (1997b)
Infectious Diseases of Coldwater Fish in Salt Water 95
105Z:\Customer\CABI\A4337 - Woo\A4419 - Woo Vouchers.vpTuesday, November 05, 2002 11:34:22 AM
Color profile: DisabledComposite Default screen

Ribosomal DNA sequence of Nucleosporasalmonis Hedrick, Groff, and Baxa, 1991(Microsporea: Enterocytozooidae): implica-tions for phylogeny and nomenclature. Jour-nal of Eukaryotic Microbiology 44, 55–60.
Drinan, E.M. and Rodger, H.D. (1990) An occur-rence of Gnathia sp., ectoparasitic isopods,on caged Atlantic salmon. Bulletin of theEuropean Association of Fish Pathologists10, 141–142.
Eaton, W.D. and Kent, M.L. (1992) A retrovirusin chinook salmon (Oncorhynchustshawytscha) with plasmacytoid leukemiaand evidence for the aetiology of the disease.Cancer Research 52, 6496–6500.
Egidius, E. (1987) Vibriosis: pathogenicity andpathology. A review. Aquaculture 67, 15–28.
Egidius, E., Andersen, K., Clausen, E. and Raa, J.(1981) Cold-water vibriosis or ‘Hitra disease’in Norwegian salmonid farming. Journal ofFish Diseases 4, 353–354.
Egidius, E. and Moster, B. (1987) Effect ofNeguvon® and Nuvan® on crabs (Cancerpagurus), lobster (Homarus americanus) andblue mussel (Mytilus edulis). Aquaculture60, 165–168.
Elston, R.A., Kent, M.L. and Harrell, L.H. (1987)An intranuclear microsporidium associatedwith acute anaemia in chinook salmon,Oncorhynchus tshawytscha. Journal ofProtozoology 34, 274–277.
Enger, Ø., Husevåg, B. and Goksøyr, J. (1989)Presence of Vibrio salmonicida in fish farmsediments. Applied Environmental Micro-biology 55, 2815–2818.
Espelid, S., Roedseth, O.M. and Jørgensen, T.O.(1991) Vaccine experiments and studies ofthe humoral immune responses in cod,Gadus morhua L., to four strains ofmonoclonal-defined Vibrio anguillarum.Journal of Fish Diseases 14, 185–197.
Evelyn, T.P.T. (1971) First records of vibriosisin Pacific salmon cultured in Canada,and taxonomic status of the responsiblebacterium, Vibrio anguillarum. Journal of theFisheries Research Board of Canada 28,517–525.
Evelyn, T.P.T. (1984) Immunization againstpathogenic vibrios. In: de Kinkelin, P. andMichel, C. (eds) Symposium on Fish Vaccina-tion. OIE Fish Diseases Commission, Paris,pp. 121–150.
Evelyn, T.P.T. (1988) Vibrio vaccines forsalmonids. In: Congress Proceedings,Aquaculture International Congress andExposition, 6–9 September 1988, Vancouver,British Columbia, Canada, pp. 459–469.
Evensen, Ø., Espelid, S. and Håstein, T.(1991a) Immunohistochemical identificationof Vibrio salmonicida in stored tissues ofAtlantic salmon, Salmo salar, from thefirst known outbreak of coldwater vibriosis(‘Hitra disease’). Diseases of Aquatic Organ-isms 10, 185–189.
Evensen, Ø., Thorud, K.E. and Olsen, Y.A. (1991b)A morphological study of the gross and lightmicroscopic lesions of infectious salmonanaemia in Atlantic salmon (Salmo salar).Research in Veterinary Science 51, 215–222.
Ferguson, H.W. and Moccia, R.D. (1980) Dissemi-nated hexamitiasis in siamese fighting fish.Journal of the American Veterinary MedicalAssociation 177, 854–857.
Ferguson, H.W., Poppe, T. and Speare, D. (1990)Cardiomyopathy in farmed Norwegiansalmon. Diseases of Aquatic Organisms 8,225–231.
Ferguson, H.W., Roberts, R.J., Richards, R.H., Col-lins, R.O. and Rice, D.A. (1986) Severe degen-erative cardiomyopathy associated with pan-creas disease in Atlantic salmon, Salmo salarL. Journal of Fish Diseases 20, 95–98.
Frelier, P., Elston, R.A., Loy, J.K. and Mincher, C.(1994) Macroscopic and microscopic fea-tures of ulcerative stomatitis in farmedAtlantic salmon Salmo salar. Diseases ofAquatic Organisms 18, 227–231.
Frerichs, G.N. and Roberts, R.J. (1989) Thebacteriology of teleosts. In: Roberts, R.J. (ed.)Fish Pathology. Baillière Tindall, London,pp. 289–319.
Fryer, J.L., Lannan, C.N., Garcés, L.H., Larenas, J.J.and Smith, P.A. (1990) Isolation of arickettsiales-like organism from diseasedcoho salmon (Oncorhynchus kisutch) inChile. Fish Pathology 25, 107–114.
Fryer, J.L., Lannan, C.N., Giovannoni, S.J. andWood, N.D. (1992) Piscirickettsia salmonisgen. nov., sp. nov., the causative agent of anepizootic disease in salmonid fishes. Inter-national Journal of Systematic Bacteriology42, 120–126.
Gaggero, A., Castro, H. and Sandino, A.M. (1995)First isolation of Piscirickettsia salmonisfrom coho salmon, Oncorhynchus kisutch(Walbaum), and rainbow trout, Oncor-hynchus mykiss (Walbaum), during thefreshwater phase of their cycle. Journal ofFish Diseases 18, 277–279.
Garcés, L.H., Larenas, J.J., Smith, P.A., Sandino,S., Lannan, C.N. and Fryer, J.L. (1991)Infectivity of a rickettsia isolated from cohosalmon Oncorhynchus kisutch. Diseases ofAquatic Organisms 11, 93–97.
96 M.L. Kent and T.T. Poppe
106Z:\Customer\CABI\A4337 - Woo\A4419 - Woo Vouchers.vpTuesday, November 05, 2002 11:34:23 AM
Color profile: DisabledComposite Default screen

Garcés, L.H., Correal, P., Larenas, J., Contreras, J.,Oyanadel, S., Fryer, J.L. and Smith-Shuster,P.A. (1994) Finding of Piscirickettsiasalmonis on Ceratothoa gaudichadii. In:Abstracts of the International Symposiumon Aquatic Animal Health, 4–8 September1994, Seattle, Washington, p. 109.
Gibson, D.I. (1996) Trematoda. In: Margolis, L. andKabata, Z. (eds) Guide to the Parasites ofFishes of Canada, Part IV. Canadian SpecialPublication of Fisheries and Aquatic Sci-ences 124, 373 pp.
González, L. and Carvajal, G.J. (1994) Parásitos enlos cultivos marinos de salmónidos en el surde Chile. Investigacion Pesquera (Chile) 38,87–96.
Grant, A.N. and Treasurer, J.W. (1993) The effectsof fallowing on caligid infestations on farmedAtlantic salmon (Salmo salar L.) in Scotland.In: Boxshall, G.A. and DeFaye, D. (eds) Patho-gens of Wild and Farmed Fish: Sea Lice. EllisHorwood, Chichester, UK, pp. 255–260.
Grant, A.N., Brown, A.G., Cox, D.I., Birkbeck,T.H. and Griffen, A.A. (1996) Rickettsia-likeorganism in farmed salmon. VeterinaryRecord 138, 423 pp. (correspondence).
Grave, K., Engelstad, M. and Sóli, N.E. (1991a)Utilization of dichlorvos and trichlorfonin salmonid farming in Norway during1981–1988. Acta Veterinaria Scandinavica32, 1–7.
Grave, K., Engelstad, M., Sóli, N.E. and Toverud,E.-L. (1991b) Clinical use of dichlorvos(Nuvan®) and trichlorfon (Neguvon®) in thetreatment of salmon louse, Lepeophtheirussalmonis: compliance with the recommen-ded treatment procedures. Acta VeterinariaScandinavica 32, 9–14.
Grayson, T.H., Jenkins, P.G., Wrathmell, A.B. andHarris, J.E. (1991) Serum responses to thesalmon louse, Lepeophtheirus salmonis(Krøyer, 1838), in naturally infectedsalmonids and immunized rainbow trout,Oncorhynchus mykiss (Walbaum), andrabbits. Fish and Shellfish Immunology 1,141–155.
Grayson, T.H., John, R.J., Wadsworth, S., Greaves,K., Cox, D., Roper, J., Wrathmell, A.B., Gilpin,M.L. and Harris, J.E. (1995) Immunization ofAtlantic salmon against the salmon louse:identification of antigens and effects on lousefecundity. Journal of Fish Biology 47 (Suppl.A), 85–94.
Grisez, L., Ceusters, R. and Ollevier, F. (1991) Theuse of API 20E for the identification of Vibrioanguillarum and V. ordalii. Journal of FishDiseases 14, 351–365.
Groman, D., Tweedie, D. and Shaw, D. (1992)Experiences with atypical furunculosis inNewfoundland: an overview. Bulletin of theAquaculture Association of Canada 92–1,36–39.
Grotmol, S., Totland, G.K., Kvellestad, A., Fjell, K.and Olsen, A.B. (1995) Mass mortality oflarval and juvenile hatchery-reared halibut(Hippoglossus hippoglossus L.) associatedwith the presence of virus-like particles invacuolated lesions in the central nervoussystem and retina. Bulletin of the EuropeanAssociation of Fish Pathologists 15, 176–180.
Grotmol, S., Totland, G.K. and Kryvi, H. (1997)Detection of a nodavirus-like agent in hearttissue from reared Atlantic salmon Salmosalar suffering from cardiac myopathy syn-drome (CMS). Diseases of Aquatic Organisms29, 79–84.
Handlinger, J., Soltani, M. and Percival, S. (1997)The pathology of Flexibacter maritimus inaquaculture. Journal of Fish Diseases 20,159–168.
Harrell, L.W. and Scott, T.M. (1985) Kudoathyrsitis (Gilchrist) (Myxosporea: Multi-valvulida) in Atlantic salmon, Salmo salar L.Journal of Fish Diseases 8, 329–332.
Harrell, L.W., Novotny, A.J., Schiewe, M.H.and Hodgins, H.O. (1976) Isolation anddescription of two vibrios pathogenic toPacific salmon in Puget Sound, Washington.Fisheries Bulletin (US) 74, 447–449.
Håstein, T. and Lindstad, T. (1991) Diseasesin wild and cultured salmon: possible inter-action. Aquaculture 98, 277–288.
Hauck, A.K. (1984) A mortality and associatedtissue reactions of chinook salmon,Oncorhynchus tshawytscha (Walbaum),caused by the microsporidan Loma sp. Jour-nal of Fish Diseases 7, 217–229.
Hedrick, R.P., Groff, J.M., McDowell, T.S., Willis,M. and Cox, W.T. (1990) Haematopoieticintranuclear microsporidian infections withfeatures of leukemia in chinook salmonOncorhynchus tshawytscha. Diseases ofAquatic Organisms 8, 189–197.
Hedrick, R.P., Groff, J.M. and Baxa, D.V. (1991a)Experimental infections with Nucleosporasalmonis n.g. n.sp.: an intranuclear micro-sporidium from chinook salmon (Oncor-hynchus tshawytscha). Fish Health Section/American Fisheries Society Newsletter 19, 5.
Hedrick, R.P., Groff, J.M. and Baxa, D.V.(1991b) Experimental infections withEnterocytozoon salmonis Chilmonczyk, Cox,Hedrick (Microsporea): an intranuclearmicrosporidium from chinook salmon
Infectious Diseases of Coldwater Fish in Salt Water 97
107Z:\Customer\CABI\A4337 - Woo\A4419 - Woo Vouchers.vpTuesday, November 05, 2002 11:34:23 AM
Color profile: DisabledComposite Default screen

Oncorhynchus tshawytscha. Diseases ofAquatic Organisms 10, 103–108.
Hervio, D.M.L., Kent, M.L., Khattra, J., Sakanari, J.,Yokoyama, H. and Devlin, R.H. (1997)Taxonomy of Kudoa species (Myxosporea:Multivalvulida) using small subunit ribo-somal DNA sequence. Canadian Journal ofZoology 75, 2112–2119.
Hicks, B. (1989) British Columbia SalmonidDisease Handbook. Province of BritishColumbia, Ministry of Agriculture andFisheries, Victoria, British Columbia, 40 pp.
Higgins, M., Dawe, S.C. and Kent, M.L. (1998) Effi-cacy of the fumagillin analog TNP-470, forNucleospora salmonis and Loma salmonaeinfections in chinook salmon Oncorhynchustshawytscha. Diseases of Aquatic Organisms34, 45–49.
Hikida, M., Wakabayashi, H., Egusa, S. andMasumura, K. (1979) Flexibacter sp., agliding bacterium pathogenic to some marinefishes in Japan. Bulletin of the Japanese Soci-ety of Scientific Fisheries 45, 421–428.
Hilger, I., Ulrich, S. and Anders, K. (1991) A newulcerative flexibacteriosis-like disease (‘yel-low pest’) affecting young Atlantic cod Gadusmorhua from the German Wadden Sea. Dis-eases of Aquatic Organisms 11, 19–29.
Hodneland, K., Nylund, A., Nilsen, F. andMidttun, B. (1993) The effect of nuvan,azamethiphos and hydrogen peroxide onsalmon lice (Lepeophtheirus salmonis).Bulletin of the European Association of FishPathologists 13, 203–206.
Hoffman, G.L. and Meyer, F.P. (1974) Parasites ofFreshwater Fish. TFH Publishers, NeptuneCity, New Jersey, 224 pp.
Holm, K.O., Strøm, E., Stensvåg, K., Raa, J.and Jørgensen, T. (1985) Characteristics of aVibrio sp. associated with the ‘Hitra Disease’of Atlantic salmon in Norwegian fish farms.Fish Pathology 20, 125–129.
Houghton, G. (1994) Acquired protection inAtlantic salmon Salmo salar parr and post-smolts against pancreas disease. Diseases ofAquatic Organisms 18, 109–118.
House, M.L., Bartholomew, J.L., Winton, J.R. andFryer, J.L. (1998) Relative virulence of threeisolates of Piscirickettsia salmonis for cohosalmon Oncorhynchus kisutch. Diseases ofAquatic Organisms 35, 107–113.
Howard, T. and Carson, J. (1993a) Verificationthat Paramoeba species are consistentlyassociated with gill damage in fish affectedwith amoebic gill disease. In: Proceedingsof the Saltas Research Review Seminar, 29April 1993, Hobart, Australia, pp. 69–80.
Howard, T. and Carson, J. (1993b) Are therealternatives to freshwater treatment of AGD?In: Proceedings of the Saltas Research ReviewSeminar, 29 April 1993, Hobart, Australia,pp. 81–87.
Inostroza, R., Sievers, G., Roa, J. and Aguirrebena,R. (1993) Prevalencia e intensidad de infecc-ion estacional por Ceratothoa gaudichaudiien salmones (Salmo salar) cutlivados en aquade mar en el sur de Chile. Archivos deMedicina Veterinaria (Valdivia) 25, 173–179.
Jarp, J., Gjevre, A.G., Olsen, A.B. and Bruheim, T.(1994) Risk factors associated with furun-culosis, infectious pancreatic necrosis andmortality in post-smolt of Atlantic salmon,Salmo salar L. Journal of Fish Diseases 18,67–78.
Jarp, J., Taksdal, T. and Trøud, B. (1996) Infectiouspancreatic necrosis in Atlantic salmon Salmosalar in relation to specific antibodies, smol-tification, and infection with erythrocyticinclusion body syndrome (EIBS). Diseases ofAquatic Organisms 27, 81–88.
Johnson, S.C. and Albright, L.J. (1991) The devel-opmental stages of Lepeophtheirus salmonis(Krøyer, 1837) (Copepoda: Caligidae). Cana-dian Journal of Zoology 69, 929–950.
Johnson, S.C. and Albright, L.J. (1992) The effectsof cortisol implants on the susceptibility andhistopathology of the responses of naive cohosalmon Oncorhynchus kisutch to experi-mental infection with Lepeophtheirussalmonis (Copepoda: Caligidae). Diseases ofAquatic Organisms 14, 195–205.
Johnson, S.C. and Margolis, L. (1993) The efficacyof ivermectin for the control of sea liceon sea-farmed Atlantic salmon. Diseasesof Aquatic Organisms 17, 107–112.
Johnson, S.C. and Margolis, L. (1994) Sea lice.In: Thoesen, J.C. (ed.) Suggested Proceduresfor the Detection and Identification ofCertain Finfish and Shellfish Pathogens, 4thedn. Fish Health Section, American FisheriesSociety, 10 pp.
Johnson, S.C., Constible, J.M. and Richard, J.(1993) Laboratory investigations of thetoxicological and histopathological effects ofhydrogen peroxide to salmon and its efficacyagainst the salmon louse Lepeophtheirussalmonis. Diseases of Aquatic Organisms 17,197–204.
Johnson, S.C., Blaylock, R.B., Elphick, J. andHyatt, K. (1996) Disease caused by the salmonlouse Lepeophtheirus salmonis (Copepoda:Caligidae) in wild sockeye salmon(Oncorhynchus nerka) stocks of AlberniInlet, British Columbia. Canadian Journal
98 M.L. Kent and T.T. Poppe
108Z:\Customer\CABI\A4337 - Woo\A4419 - Woo Vouchers.vpTuesday, November 05, 2002 11:34:23 AM
Color profile: DisabledComposite Default screen

of Fisheries and Aquatic Sciences 53,2888–2897.
Jones, M.W., Sommerville, C. and Wootten, R.(1992) Reduced sensitivity of the salmonlouse, Lepeophtheirus salmonis, to theorganophosphate dichlorvos. Journal of FishDiseases 15, 197–202.
Jones, S.R.M., MacKinnon, A.M. and Groman,D.B. (1999) Virulence and pathogenicity ofinfectious salmon anemia virus isolated fromfarmed salmon in Atlantic Canada. Journal ofAquatic Animal Health 11, 400–405.
Jónsdóttir, H., Bron, J.E., Wootten, R. andTurnbull, J.F. (1992) The histopathologyassociated with the pre-adult and adult stagesof Lepeophtheirus salmonis on the Atlanticsalmon, Salmo salar L. Journal of FishDiseases 15, 521–527.
Jørgensen, T., Midling, K., Espelid, S., Nilsen, R.and Stensvåg, K. (1989) Vibrio salmonicida,previously registered as a pathogen insalmonids, does also cause mortality innetpen captured cod (Gadus morhua).Bulletin of the European Association of FishPathologists 9, 42–44.
Kabata, Z. (1970) Crustacea as Enemies of Fishes.In: Snieszko, S.F. and Axelrod, H.R. (serieseds) Diseases of Fish. TFH Publications,Jersey City, New Jersey.
Kabata, Z. (1979) Parasitic Copepoda of BritishFishes. The Ray Society, London, 468 pp.
Kabata, Z. (1984) Diseases caused by metazoans:crustaceans. In: Kinne, O. (ed.) Diseases ofMarine Animals, Vol. 4, Part 1. BiologischeAnstalt Helgoland Hamburg, pp. 321–399.
Kabata, Z. (1988) Copepoda and Branchiura.In: Margolis, L. and Kabata, Z. (eds) Guide tothe Parasites of Fishes of Canada. Part II.Crustacea. Canadian Special Publicationof Fisheries and Aquatic Sciences 101,pp. 3–127.
Kabata, Z. and Whitaker, D.J. (1981) Two speciesof Kudoa (Myxosporea: Multivalvulida)parasitic in the flesh of Merluccius productus(Ayres, 1855) (Pisces: Teleostei) in theCanadian Pacific. Canadian Journal ofZoology 59, 2085–2091.
Kent, M.L. (1982) Characteristics and identifica-tion of Pasteurella and Vibrio species patho-genic to fishes using API-20E (AnalytabProducts) Multitube Test Strips. CanadianJournal of Fisheries and Aquatic Sciences 39,1725–1729.
Kent, M.L. and Dawe, S.C. (1990) Transmissionof a plasmacytoid leukemia of chinooksalmon Oncorhynchus tshawytscha. CancerResearch (Suppl.) 50, 5679s–5681s.
Kent, M.L. and Dawe, S.C. (1994) Efficacy ofFumagillin DCH against experimentally-induced Loma salmonae (Microsporea)infections in chinook salmon Oncorhynchustshawytscha. Diseases of Aquatic Organisms20, 231–233.
Kent, M.L. and Elston, R.A. (1987) Diseases ofseawater reared salmon in Washington State.In: Proceedings of the Fourth AlaskaAquaculture Conference. Alaska Sea GrantReport No. 88 4, pp. 161–163.
Kent, M.L. and Poppe, T.T. (1998) Diseases ofSeawater Netpen-reared Salmonids. PacificBiological Station Press, Nanaimo, BritishColumbia, Canada, 138 pp.
Kent, M.L., Dungan, C.F., Elston, R.A. and Holt,R.A. (1988a) Cytophaga sp. (Cytophagales)infection in seawater pen-reared Atlanticsalmon Salmo salar. Diseases of AquaticOrganisms 4, 173–179.
Kent, M.L., Sawyer, T.K. and Hedrick, R.P.(1988b) Paramoeba pemaquidensis (Sarco-mastigophora: Paramoebidae) infestation ofthe gills of coho salmon Oncorhynchuskisutch reared in seawater. Diseases ofAquatic Organisms 5, 163–169.
Kent, M.L., Elliot, D.G., Groff, J.M. and Hedrick,R.P. (1989) Loma salmonae (Protozoa:Microspora) infections in seawater rearedcoho salmon (Oncorhynchus kisutch).Aquaculture 80, 211–222.
Kent, M.L., Groff, J.M., Traxler, G.S., Zinkl, J.G.and Bagshaw, J.W. (1990) Plasmacytoidleukemia in seawater reared chinook salmonOncorhynchus tshawytscha. Diseases ofAquatic Organisms 8, 199–209.
Kent, M.L., Margolis, L. and Fournie, J.W. (1991)A new eye disease in pen-reared chinookcaused by metacestodes of Gilquinia squali(Trypanorhyncha). Journal of Aquatic Ani-mal Health 3, 134–140.
Kent, M.L., Ellis, J., Fournie, J.W., Dawe, S.C.,Bagshaw, J.W. and Whitaker, D. (1992)Systemic hexamitid infection in seawaterpen-reared chinook salmon Oncorhynchustshawytscha. Diseases of Aquatic Organisms14, 81–89.
Kent, M.L., Dawe, S.C. and Speare, D.J. (1995)Transmission of Loma salmonae (Micro-sporea) to chinook salmon in seawater.Canadian Veterinary Journal 36, 98–102.
Kent, M.L., Whitaker, D.J., Moran, J. and Kabata, Z.(1997) Haemobaphes disphaerocephalus(Pennellidae: Copepoda), an accidentalparasite of seawater pen-reared Atlanticsalmon. Canadian Veterinary Journal 38,110–111.
Infectious Diseases of Coldwater Fish in Salt Water 99
109Z:\Customer\CABI\A4337 - Woo\A4419 - Woo Vouchers.vpTuesday, November 05, 2002 11:34:23 AM
Color profile: DisabledComposite Default screen

Kent, M.L., Traxler, G.S., Kieser, D., Richard, J.,Dawe, S.C., Shaw, R.W., Prosperi-Porta, G.,Ketcheson, J. and Evelyn, T.P.T. (1998) Sur-vey of salmonid pathogens in ocean-caughtfishes in British Columbia, Canada. Journalof Aquatic Animal Health 10, 211–219.
Khalil, L.F., Jones, A. and Bray, R.A. (eds) (1994)Keys to the Cestode Parasites of Vertebrates.CAB International, Wallingford, UK, 751 pp.
Khan, R.A. (1988) Experimental transmission,development, and effects of a parasiticcopepod, Lernaeocera branchialis, onAtlantic cod, Gadus morhua. Journal ofParasitology 74, 586–599.
Khan, R.A., Lee, E.M. and Barker, D. (1990)Lernaeocera branchialis: a potential patho-gen to cod ranching. Journal of Parasitology76, 913–917.
Kimura, T. and Yoshimizu, M. (1991) Viraldiseases of fish in Japan. Annual Reviewof Fish Diseases 1, 67–82.
Kimura, T., Yoshimizu, M. and Tanaka, M. (1981a)Studies on a new virus (OMV) from Oncor-hynchus masou. II. Oncogenic nature. FishPathology 15, 149–153.
Kimura, T., Yoshimizu, M., Tanaka, M. andSannohe, H. (1981b) Studies on a newvirus (OMV) from Oncorhynchus masou.I. Characteristics and pathogenicity. FishPathology 15, 149–153.
Kimura, T., Suzuki, S. and Yoshimizu, M.(1983) In vitro antiviral effect of 9-(2hydroxy-ethoxymethyl) guanine on the fish herpesvirus (OMV). Antiviral Research 3, 103–108.
Kitao, I., Aoki, I. and Muroga, K. (1984) Three newO-serotypes of Vibrio anguillarum. Bulletinof the Japanese Society of Scientific Fisheries50, 1955.
Krogsrud, J., Håstein, T. and Ronningen, K. (1989)Infectious pancreatic necrosis virus inNorwegian fish farms. In: Ahne, W. andKurstak, E. (eds) Viruses of Lower Verte-brates. Springer-Verlag, Berlin, pp. 284–291.
Kumagai, A., Takahashi, K. and Fukuda, H. (1994)Epizootics caused by salmonid herpesvirustype 2 infection in maricultured cohosalmon. Fish Pathology 29, 127–134.
Kumagai, A., Takahashi, K. and Fukuda, H. (1997)Infection source of herpesvirus disease incoho salmon culture and its control. FishPathology 32, 103–108.
Kvenseth, P.G. (1993) Use of wrasse to controlsea lice. In: Reinersten, H., Dahle, L.A.,Jørgensen, T.L. and Tvinnereim, K. (eds) FishFarming Technology. Balkema, Rotterdam,pp. 227–232.
Lannan, C.N. and Fryer, J.L. (1991) Recommendedmethods for inspection of fish for thesalmonid ricketttsia. Bulletin of the Euro-pean Association of Fish Pathologists 11,135–136.
Lannan, C.N. and Fryer, J.L. (1994) Extracellularsurvival of Piscirickettsia salmonis. Journalof Fish Diseases 17, 545–548.
Lannan, C.N., Ewing, S.A. and Fryer, J.L. (1991)A fluorescent antibody test for detectionof the rickettsia causing disease in Chileansalmonids. Journal of Aquatic Animal Health3, 229–234.
Larenas, J.J., Astroga, C., Contreras, J. and Smith, P.(1996) Detección de Piscirickettsia salmonisen ovas fertilizadas provenientes de truchaarco iris (Oncorhynchus mykiss) experimen-talemente infectadas. Archivo di MedicinaVeterinaria (Valdivia) 28, CommunicacionesNo. 2.
Lysne, D.A., Hemmingsen, W. and Skorping, A.(1994) The distribution of Cryptocotyle spp.metacercariae in the skin of caged Atlanticcod (Gadus morhua). Journal of Fish Biology45, 352–355.
MacKenzie, K. (1965) The plerocercoid ofGilquinia squali Fabricius, 1794. Parasit-ology 55, 607–615.
MacKenzie, K. (1975) Some aspects of the biologyof the plerocercoid of Gilquinia squaliFabricius 1794 (Cestoda: Trypanorhyncha).Journal of Fish Biology 7, 321–327.
MacKinnon, B.M. (1991) Sea lice and Atlanticsalmon: absence of immunoprotection inSalmo salar to Caligus elongatus. Bulletin ofthe Aquaculture Association of Canada 91,58–60.
Markey, P.T., Blazer, V.S., Ewing, M.S. and Kocan,K.M. (1994) Loma sp. in salmonids fromeastern United States: associated lesions inrainbow trout. Journal of Aquatic AnimalHealth 6, 318–328.
Masumura, K. and Wakabayashi, H. (1977) Anoutbreak of gliding bacterial disease inhatchery-born red seabream (Pagrus major)and gilthead (Acanthopagrus schlegeli) fry inHiroshima. Fish Pathology 12, 171–177.
Mattis, T.E. (1986) Development of two tetrarhyn-chidean cestodes from the northern Gulfof Mexico. PhD dissertation. University ofSouthern Mississippi, Mississippi, 172 pp.
Mauel, M.J., Giovannoni, S.J. and Fryer, J.L. (1996)Development of polymerase chain reactionassays for detection, identification, anddifferentiation of Piscirickettsia salmonis.Diseases of Aquatic Organisms 26, 189–195.
100 M.L. Kent and T.T. Poppe
110Z:\Customer\CABI\A4337 - Woo\A4419 - Woo Vouchers.vpTuesday, November 05, 2002 11:34:23 AM
Color profile: DisabledComposite Default screen

McElwain, I.B. and Post, G. (1968) Efficacyof cyzine for trout hexamitiasis. The Pro-gressive Fish-Culturist 30, 84–91.
McGladdery, S.E., Murphy, L., Hicks, B.D. andWagner, S.K. (1990) The effects of Stephano-stomum tenue (Digenea: Acanthocolpidae)on marine aquaculture of rainbow trout,Salmo gairdneri. In: Perkins, F.O. and Cheng,T.C. (eds) Pathology in Marine Science.Academic Press, San Diego, California,pp. 305–315.
McLoughlin, M.F., Nelson, R.T., Rowley, H.M.,Cox, D.I. and Grant, A.N. (1996) Experi-mental pancreas disease in Atlantic salmonSalmo salar post-smolts induced by salmonpancreas disease virus (SPDV). Diseases ofAquatic Organisms 26, 117–124.
McVicar, A.H. (1987) Pancreas disease of farmedAtlantic salmon, Salmo salar, in Scotland:epidemiology and early pathology. Aqua-culture 67, 71–78.
McVicar, A.H. and Munro, A.L.S. (1987) PancreasDisease in Farmed Atlantic Salmon: Evi-dence for an Infectious Aetiology. Inter-national Council for the Exploration of theSea – CM 1987/F:6 Mariculture Committee.
Melby, H.P., Krogsrud, J., Håstein, T. and Stenwig,H. (1991) All commercial Atlantic salmonseawater farms in Norway harbour carriers ofinfectious pancreatic necrosis virus (IPNV).In: Fryer, J.L. (ed.) Proceedings of the 2ndInternational Symposium on Viruses ofLower Vertebrates. Oregon State University,Corvallis, Oregon, pp. 211–217.
Menzies, F.D., Wheatley, S.B., Goodall, E.A. andMcLoughlin, M.F. (1996) Development of acomputerised information retrieval systemfor Atlantic salmon (Salmo salar). Aqua-culture Research 27, 183–190.
Mitchell, C.G. (1993) Eubothrium. AquacultureInformation Series 14, Scottish OfficeAgriculture and Fisheries for Scotland,Aberdeen, UK.
Mjaaland, S., Rimstad, E., Falk, K. and Dannevig,B.H. (1997) Genomic characterization of thevirus causing infectious salmon anaemiain Atlantic salmon (Salmo salar L.): anorthomyxo-like virus in a teleost. Journal ofVirology 71, 7681–7686.
Mo, T.A., Poppe, T.T. and Iversen, L. (1990)Systemic hexamitosis in salt-water rearedAtlantic salmon. Bulletin of the EuropeanAssociation of Fish Pathologists 10, 69–70.
Möller, H. and Anders, K. (1986) Diseases andParasites of Marine Fishes. Verlag Möller,Kiel, Germany, 365 pp.
Moran, J.D.W., Margolis, L., Webster, J.M. andKent, M.L. (1999a) Sequential developmentof Kudoa thyrsites (Myxozoe: Myxosporea) innetpen-reared Atlantic salmon determinedby light microscopy and a polymerase chainreaction test.. Diseases of Aquatic Organisms37, 185–193.
Moran, J.D.W., Whitaker, D.J. and Kent, M.L.(1999b) Natural and laboratory transmissionof the marine myxosporean parasite Kudoathyrsites to Atlantic salmon. Journal ofAquatic Animal Health 11, 110–115.
Morrison, C.M. (1983) The distribution of themicrosporidian Loma morhua in tissues ofthe cod Gadus morhua L. Canadian Journalof Zoology 61, 2155–2161.
Morrison, C.M. and Sprague, V. (1981) Micro-sporidian parasites in the gills of salmonidfishes. Journal of Fish Diseases 4, 371–386.
Morrison, C.M. and Sprague, V. (1983) Lomasalmonae (Putz, Hoffman and Dunbar 1965)in the rainbow trout, Salmo gairdneriRichardson, and L. fontinalis sp. nov.(Microsporida) in the brook trout, Salvelinusfontinalis (Mitchell). Journal of Fish Diseases6, 345–353.
Morrison, J.K., MacConnell, E., Chapman, P.F. andWestgard, R.L. (1990) A microsporidium-induced lymphoblastosis in chinook salmonOncorhynchus tshawytscha. Diseases ofAquatic Organisms 8, 99–104.
Mullins, J.E., Powell, M., Speare, D.J. andCawthorn, R. (1994) An intranuclear micro-sporidian in lumpfish Cyclopterus lumpus.Diseases of Aquatic Organisms 20, 7–13.
Munday, B.L., Lange, K., Foster, C., Lester, R.J.G.and Handlinger, J. (1993) Amoebic gill dis-ease of sea-caged salmonids in Tasmanianwaters. Tasmanian Fisheries Research 28,14–19.
Mundry, D.R. and Dailey, M.D. (1971)Postembryonic development of certaintetraphyllidean and trypanorhynchancestodes with a possible alternative lifecycle for the order Trypanorhyncha. Cana-dian Journal of Zoology 49, 1249–1253.
Munro, A.L.S., Ellis, A.E., McVicar, A.H., McLay,A.H. and Needham, E.A. (1984) An exocrinepancreas disease of farmed Atlantic salmonin Scotland. Helgolaender Meeresunterssu-chungen 37, 571–586.
Nagasawa, K. and Sakamoto, S. (1993) Infection ofthe salmon louse Lepeophtheirus salmonis(Copepoda: Caligidae) on seawater-farmedsalmonids in Northern Honshu, Japan.Grobyo Kenkyu 28, 143–144.
Infectious Diseases of Coldwater Fish in Salt Water 101
111Z:\Customer\CABI\A4337 - Woo\A4419 - Woo Vouchers.vpTuesday, November 05, 2002 11:34:24 AM
Color profile: DisabledComposite Default screen

Nelson, R.T., McLouglin, M.F., Rowley, H.M.,Platten, M.A. and McCormick, J.I. (1995)Isolation of a toga-like virus from farmedAtlantic salmon Salmo salar with pancreasdisease. Diseases of Aquatic Organisms 22,25–32.
Newbound, G.C. and Kent, M.L. (1991)Experimental interspecies transmission ofplasmacytoid leukemia in salmonid fishes.Diseases of Aquatic Organisms 10, 159–166.
Nilsen, F., Ness, A. and Nylund, A. (1995) Obser-vations on an intranuclear microsporidian inlymphoblasts from farmed Atlantic halibutlarvae (Hippoglossus hippoglossus L.). Jour-nal of Eukaryotic Microbiology 42, 131–135.
Novotny, A.J. (1978) Vibriosis and furunculosisin marine cultured salmon in Puget Sound,Washington. Marine Fisheries Review 40,52–55.
Novotny, A.J. and Mahnken, C.V.W. (1971)Predation on juvenile Pacific salmon by amarine isopod Rocinela belliceps pugettensis(Crustacea, Isopoda). US National MarineFisheries Service Fishery Bulletin 69,699–701.
Nylund, A. and Jakobsen, P. (1995) Sea trout as acarrier of infectious salmon anaemia virus.Journal of Fish Biology 47, 174–176.
Nylund, A., Hovland, T., Hodneland, K., Nilsen, F.and Løvik, P. (1994) Mechanisms for trans-mission of infectious salmon anaemia (ISA).Diseases of Aquatic Organisms 19, 95–100.
O’Halloran, J. and Henry, R. (1993) Vibriosalmonicida (Hitra disease) in New Bruns-wick. Bulletin of the Aquaculture Associa-tion of Canada 4, 96–98.
Olsen, A.B., Melby, H.P., Speilberg, L., Eversen,Ø. and Håstein, T. (1997) Piscirickettsiasalmonis infection in Atlantic salmon Salmosalar in Norway – epidemiological, patholog-ical and microbiological findings. Diseases ofAquatic Organisms 31, 35–48.
Overstreet, R.M. (1978) Marine Maladies? Worms,Germs and Other Symbionts From the North-ern Gulf of Mexico. Blossman Printing, OceanSprings, Mississippi, 140 pp.
Pacha, R.E. and Ordal, E.J. (1970) Myxobacterialdiseases of salmonids. In: Snieszko, S.F. (ed.)A Symposium on Diseases of Fishes andShellfishes. Special Publication No.5,American Fisheries Society, Washington,DC, pp. 243–247.
Palmer, R. (1994) Kudoa – the Irish experience.In: Conley, D.C. (ed.) Kudoa Workshop Pro-ceedings, 17–18 February 1994, Nanaimo,British Columbia. Aquaculture IndustryDevelopment Report 94-01: 18-21, Province
of British Columbia, Ministry of Agricul-ture, Fisheries and Food, Victoria, BritishColumbia, Canada.
Palmer, R., Rodger, H., Drian, E., Dwyer, C.and Smith, P.R. (1987) Preliminary trials onthe efficacy of Ivermectin against parasiticcopepods of Atlantic salmon. Bulletin of theEuropean Association of Fish Pathologists 7,47–54.
Patashnik, M., Groninger, H.S., Barnett, H.,Kudo, G. and Koury, B. (1982) Pacificwhiting, Merluccius productus. I. Abnormalmuscle texture caused by myxosporidian-induced proteolysis. Marine FisheriesReview 44, 1–12.
Poppe, T.T. and Mo, T.A. (1993) Systemic,granulomatous hexamitosis of farmedAtlantic salmon: interaction with wild fish.Fisheries Research 17, 147–152.
Poppe, T.T., Mo, T.A. and Iversen, L. (1992)Disseminated hexamitosis in sea-cagedAtlantic salmon Salmo salar. Diseases ofAquatic Organisms 14, 91–97.
Poynton, S.L. (1986) Distribution of the flagellateHexamita salmonis Moore, 1922 and themicrosporidian Loma salmonae Putz,Hoffman and Dunbar, 1965 in brown trout,Salmo trutta L., and rainbow trout, Salmogairdneri Richardson, in the River Itchen(UK) and three of its fish farms. Journal ofFish Biology 29, 417–429.
Putz, R.E. and McLaughlin, J.J.A. (1970) Thebiology of Nosematidae (Microsporida)from freshwater and euryhaline fishes.In: Snieszko, S.F. (ed.) A Symposium onDiseases of Fishes and Shellfishes. SpecialPublication No. 5, American Fisheries Soci-ety, Washington, DC, pp. 124–132.
Putz, R.E., Hoffman, G.L. and Dunbar, C.E. (1965)Two new species of Plistophora (Micro-sporidea) from North American fish with asynopsis of Microsporidea of freshwater andeuryhaline fishes. Journal of Protozoology 12,228–236.
Reichenbach, H. (1988) Order Cytophagales.In: Holt, J.G., Staley, J.T., Bryant, M.P.and Pfennig, N. (eds) Bergey’s Manualof Systematic Bacteriology, Vol. 3,Williams & Wilkins, Baltimore, Maryland,pp. 2011–2066.
Rodger, H.D. and Drinan, E.M. (1993) Observa-tions on a rickettsia-like organism in Atlanticsalmon, Salmo salar L., in Ireland. Journal ofFish Diseases 16, 361–369.
Rodger, H.D. and McArdle, J.F. (1996) An outbreakof amoebic gill disease in Ireland. TheVeterinary Record 139, 348–349.
102 M.L. Kent and T.T. Poppe
112Z:\Customer\CABI\A4337 - Woo\A4419 - Woo Vouchers.vpTuesday, November 05, 2002 11:34:24 AM
Color profile: DisabledComposite Default screen

Roth, M., Richards, R.H. and Sommerville, C.(1993a) Current practices in the chemo-therapeutic control of sea lice infestationsin aquaculture: a review. Journal of FishDiseases 16, 1–21.
Roth, M., Richards, R.H. and Sommerville, C.(1993b) Preliminary studies on the efficacy oftwo pyrethroid compounds, resmethrin andlambdacyhalothrin, for the treatment of sealice (Lepeophtheirus salmonis) infestationsof farmed Atlantic salmon (Salmo salar). In:Boxshall, G.A. and DeFaye, D. (eds) Patho-gens of Wild and Farmed Fish: Sea Lice. EllisHorwood, Chichester, UK, pp. 275–289.
Roth, M., Richards, R.H., Dobson, D.P. andRae, G.H. (1996) Field trials on the efficacyof the organophosphorous compoundazamethiphos for the control of sea lice(Copepoda: Caligidae) infestations of farmedAtlantic salmon (Salmo salar). Aquaculture140, 197–217.
Roubal, F.R., Lester, R.J.G. and Foster, C.K. (1989)Studies on cultured and gill-attachedParamoeba sp. (Gymnamoebae: Paramoe-bidae) and the cytopathology of paramoebicgill disease in Atlantic salmon, Salmo salarL., from Tasmania. Journal of Fish Diseases12, 481–492.
Rucker, R.R. (1963) Status of fish diseasesand relation to production. In: Report ofthe 2nd Governor’s Conference on PacificSalmon, January 1963, Seattle, Washington,pp. 98–101.
Sakanari, J.A. and Moser, M. (1989) Completelife cycle of the elasmobranch cestode,Lacistorhynchus dollfusi Beveridge andSakanari, 1987 (Trypanorhyncha). Journal ofParasitology 75, 806–808.
Sande, R.D. and Poppe, T.T. (1995) Diagnosticultrasound examination and echocardio-graphy in Atlantic salmon (Salmo salar).Veterinary Radiology and Ultrasound 36,551–558.
Sano, T., Fukuda, H., Okamoto, N. and Kaneko, F.(1983) Yamame tumor virus: lethalityand oncogenicity. Bulletin of the JapaneseSociety of Scientific Fisheries 49, 1159–1163.
Sawyer, E.S. (1976) An outbreak of myxobacterialdisease in coho salmon (Oncorhynchuskisutch) reared in a Maine estuary. Journal ofWildlife Diseases 12, 575–578.
Scalati, F. and Kusuda, R. (1986) Chemicalcomposition of the lipopolysaccharides fromVibrio anguillarum and Vibrio ordalii. FishPathology 21, 193–199.
Schiewe, M.H., Trust, T.J. and Crosa, J.H. (1981)Vibrio ordalii sp. nov.: a causative agent
of vibriosis in fish. Current Microbiology6, 343–348.
Schmahl, G., Taraschewski, H. and Mehlhorn,H. (1989) Chemotherapy of fish parasites.Parasitology Research 75, 503–511.
Schmidt, G.D. (1986) Handbook of TapewormIdentification. CRC Press, Boca Raton,Florida, 675 pp.
Schram, T.A. (1993) Supplementary descriptionsof the developmental stages of Lepeoph-theirus salmonis (Krøyer, 1837) (Copepoda:Caligidae). In: Boxshall, G.A. and DeFaye, D.(eds) Pathogens of Wild and Farmed Fish:Sea Lice. Ellis Horwood, Chichester, UK,pp. 30–47.
Schroder, M.B., Espelid, S. and Jørgensen, T.O.(1992) Two serotypes of Vibrio salmonicidaisolated from diseased cod (Gadus morhuaL.): virulence, immunological studies andvaccination experiments. Fish and ShellfishImmunology 2, 211–221.
Seymour, T.A., Morrissey, M.T., Peters, M.Y.and An, H. (1994) Purification and character-ization of Pacific whiting proteases. Journalof Agricultural and Food Chemistry 42,2421–2427.
Shaw, R.W., Kent, M.L., Docker, M.F., Devlin,R.H., Brown, A. and Adamson, M.L. (1997) Anew Loma species (Microsporea) in shinerperch (Cymatogaster aggregata). Journal ofParasitology 83, 296–301.
Shaw, R.W., Kent, M.L. and Adamson, M.L. (1999)Iodophor treatment is not completelyefficacious in preventing Loma salmonae(Microsporidia) transmission in experi-mentally challenged chinook salmon,Oncorhynchus tshawytscha (Walbaum).Journal of Fish Diseases 22, 1–3.
Shaw, R.W., Kent, M.L. and Adamson, M.L. (2000)Innate susceptibility differences in chinooksalmon Oncorhynchus tshawytscha to Lomasalmonae (Micrsporidia). Diseases ofAquatic Organisms 43, 49–53.
Sievers, G., Palacios, P., Inostroza, R. andDolz, H. (1995) Evaluation of the toxicityof 8 insecticides in Salmo salar and thein vitro effects against the isopod parasite,Ceratothoa gaudichaudii. Aquaculture 143,9–16.
Smail, D.A., Bruno, D.W., Dear, G., McFarlane, L.and Ross, K. (1992) Infectious pancreaticnecrosis (IPN) virus Sp serotype in farmedAtlantic salmon post-smolts associated withmortality and clinical disease. Journal of FishDiseases 15, 77–83.
Smail, D.A., McFarlane, L., Bruno, D.W. andMcVicar, A.H. (1995) The pathology of an
Infectious Diseases of Coldwater Fish in Salt Water 103
113Z:\Customer\CABI\A4337 - Woo\A4419 - Woo Vouchers.vpTuesday, November 05, 2002 11:34:24 AM
Color profile: DisabledComposite Default screen

IPN-Sp subtype (Sh) in farmed Atlanticsalmon, Salmo salar in the Shetland Isles,Scotland. Journal of Fish Diseases 18,631–638.
Smith, P.R., Moloney, M., McElligott, A., Clarke,S., Palmer, R., O’Kelly, J. and O’Brien, J.(1993) The efficacy of oral ivermectin in thecontrol of sea lice infections of farmed Atlan-tic salmon. In: Boxshall, G.A. and DeFaye, D.(eds) Pathogens of Wild and Farmed Fish:Sea Lice. Ellis Horwood, Chichester, UK,pp. 296–307.
Sorensen, U.B.S. and Larsen, J.L. (1986) Sero-typing of Vibrio anguillarum. Applied andEnvironmental Microbiology 51, 593–597.
Speare, D.J., Brackett, J. and Ferguson, H.W. (1989)Sequential pathology of the gills of cohosalmon with a combined diatom andmicrosporidian gill infection. CanadianVeterinary Journal 30, 571–575.
Speare, D.J., Beaman, H.J., Jones, S.R.M.,Markham, R.J.F. and Arsenault, G.J. (1998)Induced resistance in rainbow trout,Oncorhynchus mykiss (Walbaum), to gilldisease associated with the microsporidiangill parasite Loma salmonae. Journal of FishDiseases 21, 93–100.
St-Hilaire, S., Hill, M., Kent, M.L., Whitaker, D.J.and Ribble, C.S. (1997a) A comparative studyof muscle texture and intensity of Kudoathyrsites infection in pen-reared Atlanticsalmon. Diseases of Aquatic Organisms 31,221–225.
St-Hilaire, S., Ribble, C., Whitaker, D.J. and Kent,M.L. (1997b) Evaluation of a nondestructivediagnostic test for Kudoa thyrsites in farmedAtlantic salmon (Salmo salar). Aquaculture156, 139–144.
St-Hilaire, S., Ribble, C., Whitaker, D.J. and Kent,M. (1998) Prevalence of Kudoa thyrsites insexually mature and immature pen-rearedAtlantic salmon (Salmo salar) in BritishColumbia, Canada. Aquaculture 162, 69–77.
St-Hilaire, S., Ribble, C., Traxler, G.S., Davies, T.and Kent, M.L. (2001) Evidence for a carrierstate of infectious hematopoietic necrosisvirus in chinook salmon (Oncorhynchustshawytascha). Diseases of Aquatic Organ-isms 46, 173–179.
Sterud, E., Mo, T.A. and Poppe, T.T. (1997)Ultrastucture of Spironucleus barkhanusn.sp. (Diplomodadida: Hexamitidae) fromgrayling Thymallus thymallus (L.)(Salmonidae) and Atlantic salmon Salmosalar L. (Salmonidae). Journal of EukaryoticMicrobiology 44, 399–407.
Summerfelt, R.C. and Warner, M.C. (1970)Incidence and intensity of infection ofPlistophora ovariae, a microsporidianparasite of golden shiner, Notemigonuscrysoleucas. In: Snieszko, S.F. (ed.) A Sympo-sium on Diseases of Fishes and Shellfishes.Special Publication No. 5, American Fish-eries Society, Washington, DC, pp. 142–160.
Svendsen, Y.S. (1990) Hosts of third stagelarvae of Hysterothylacium sp. (Nematoda:Anisakidae) in zooplankton from outerOslofjord, Norway. Sarsia 75, 161–167.
Tajima, K., Ezura, Y. and Kimura, T. (1985)Studies on the taxonomy and serology ofcausative organisms of fish vibriosis. FishPathology 20, 131–142.
Thomassen, J.M. (1993a) Hydrogen peroxide asa delousing agent for Atlantic salmon. In:Boxshall, G.A. and DeFaye, D. (eds) Patho-gens of Wild and Farmed Fish: Sea Lice. EllisHorwood, Chichester, UK, pp. 290–295.
Thomassen, J.M. (1993b) A new method for thecontrol of salmon lice. In: Reinersten, H.,Dahle, L.A., Jørgensen, L. and Tvinnereim, K.(eds) Fish Farming Technology. Balkema,Rotterdam, pp. 233–236.
Thorud, K. and Djupvik, H.O. (1988) Infectiousanaemia in Atlantic salmon (Salmo salar L.).Bulletin of the European Association of FishPathologists 8, 109–111.
Totland, G.K., Hjeltnes, B.K. and Flood, P.R.(1996) Transmission of infectious salmonanaemia (ISA) through natural secretions andexcretions from infected smolts of Atlanticsalmon Salmo salar during their presympto-matic phase. Diseases of Aquatic Organisms26, 25–31.
Traxler, G.S. and Richard, J. (1996) First detectionof infectious haematopoietic necrosis virusin marine fishes. Fish Health Section/American Fisheries Society Newsletter 24, 7.
Traxler, G.S., Roome, J.R. and Kent, M.L. (1993)Transmission of infectious hematopoieticnecrosis virus in seawater. Diseases ofAquatic Organisms 16, 111–114.
Treasurer, J.W. (1993) Management of sea lice(Caligidae) with wrasse (Labridae) onAtlantic salmon (Salmo salar L.) farms. In:Boxshall, G.A. and DeFaye, D. (eds) Patho-gens of Wild and Farmed Fish: Sea Lice. EllisHorwood, Chichester, UK, pp. 335–345.
Tsuyuki, H., Williscroft, S.N., Kabata, Z. andWhitaker, D.J. (1982) The relationshipbetween acid and neutral protease activitiesand the incidence of soft cooked texture inthe muscle tissue of Pacific hake Merluccius
104 M.L. Kent and T.T. Poppe
114Z:\Customer\CABI\A4337 - Woo\A4419 - Woo Vouchers.vpTuesday, November 05, 2002 11:34:24 AM
Color profile: DisabledComposite Default screen

productus infected with Kudoa paniformisand/or K. thyrsites, and held for varying timesunder different pre-freeze chilled storageconditions. Canadian Technical Report ofFisheries and Aquatic Sciences 1130, 39 pp.
Tully, O., Poole, W.R., Whelan, K.F. andMerigoux, S. (1993) Parameters and possiblecauses of epizootics of Lepeophtheirussalmonis (Krøyer) infesting sea trout (Salmotrutta L.) off the west coast of Ireland. In:Boxshall, G.A. and DeFaye, D. (eds) Patho-gens of Wild and Farmed Fish: Sea Lice. EllisHorwood, Chichester, UK, pp. 202–213.
Tully, O., Daly, P., Lysaght, S., Deady, S. andVarian, S.J.A. (1996) Use of cleaner-wrasse(Ctenolabrus exoletus (L.) and Ctenolabrusrupestris (L.)) to control infestation of Caliguselongatus Nordmann on farmed Atlanticsalmon. Aquaculture 142, 11–24.
Urawa, S. and Kato, T. (1991) Heavy infections ofCaligus orientalis (Copepoda: Caligidae) oncaged rainbow trout Oncorhynchus mykiss inbrackish water. Gyobyo Kenkyu 26, 161–162.
Vaney, C. and Conte, A. (1901) Sur une nouvellemicrosporidie, Pleistophora mirandellae,parasite de l’ovaire d’Alburnus mirandellaBlanch. Comptes Rendus de l’Academie desSciences 133, 644–646.
Vismanis, K.O., Lullu, A.V., Inglis, V.A.,Turovskii, A.M. and Yun, A.I. (1984)Diseases of Salmonidae in the marine fishfarms on the Baltic coast and their preven-tion. In: Bauer, O.N., Musselius, V.A. andSkryabina, E.S. (eds) Biologicheskie OsnovyRybovodstva: Parazity i Bolezni Ryb, ‘Nauka’,Moscow, pp. 56–63 (in Russian).
Wakabayashi, H., Hikida, M. and Masumura, K.(1984) Flexibacter infection in culturedmarine fish in Japan. Helgolaender Meere-suntersuchungen 38, 1–8.
Wakabayashi, H., Hikida, M. and Masumura, K.(1986) Flexibacter maritimus sp. nov., apathogen of marine fishes. InternationalJournal of Systematic Bacteriology 36,396–398.
Wales, J.H. and Wolf, H. (1955) Three protozoandiseases of trout in California. California Fishand Game 41, 183–187.
Wards, B.J., Patel, H.H., Andersen, C.D. andde Lisle, G.W. (1991) Characterization byrestriction endonuclease analysis andplasmid profiling of Vibrio ordalii strainsfrom salmon (Oncorhynchus tshawytschaand Oncorhynchus nerka) with vibriosisin New Zealand. New Zealand Journalof Marine and Freshwater Research 25,345–350.
Whitaker, D.J. and Kent, M.L. (1991) Kudoathrysites (Myxosporea): a cause of soft fleshin farm-reared Atlantic salmon (Salmo salar).Journal of Aquatic Animal Health 3,291–294.
Whitaker, D.J. and Kent, M.L. (1992) Kudoathyrsites (Myxosporea) and soft flesh in pen-reared coho salmon. Fish Health Section/American Fisheries Society Newsletter 20,4–5.
Whitaker, D.J., Kent, M.L. and Margolis, L. (1994)Myxosporean parasites and their potential.impact on the aquaculture industry, withemphasis on Kudoa species. In: Conley,D.C. (ed.) Kudoa Workshop Proceedings,17–18 February 1994, Nanaimo, BritishColumbia. Aquaculture Industry Develop-ment Report 94-01, Province of BritishColumbia, Ministry of Agriculture, Fisheriesand Food, Victoria, British Columbiapp. 2–7.
Williams, H. and Jones, A. (1994) Parasitic Wormsof Fish. Taylor & Francis, London, 593 pp.
Wolf, K. (1988) Fish Viruses and Fish ViralDiseases. Cornell University Press, Ithaca,New York, 476 pp.
Woo, P.T.K. and Poynton, S.L. (1995) Diplo-monadida, Kinetoplastida and Amoebida(phylum Sarcomastigophora). In: Woo,P.T.K. (ed.) Fish Diseases and Disorders,Vol. 1. Protozoan and Metazoan Infections.CAB International, Wallingford, UK, pp.27–96.
Wood, B.P., Bruno, D.W. and Ross, K. (1996)Infectious pancreatic necrosis virus (IPNV)mortalities among farmed Atlantic halibut,Hippoglossus hippoglossus L., in Scotland.Bulletin of the European Association of FishPathologists 16, 214–216.
Wood, J. (1974) Diseases of Pacific Salmon: TheirPrevention and Treatment. WashingtonState Department of Fisheries, Olympia,Washington, 82 pp.
Wootten, R., Smith, J.W. and Needham, E.A.(1982) Aspects of the biology of the parasiticcopepods Lepeophtheirus salmonis andCaligus elongatus on farmed salmonids, andtheir treatment. Proceedings of the RoyalSociety of Edinburgh, Section B (BiologicalSciences) 81, 185–197.
Yasutake, W.T., Buhler, D.R. and Shanks, W.E.(1961) Chemotherapy of hexamitiasis in fish.Journal of Parasitology 47, 81–86.
Yoshimizu, M., Fukuda, H., Sano, T. andKimura, T. (1995) Salmonid herpevirus 2.Epizootiology and serological relationship.Veterinary Research 26, 486–492.
Infectious Diseases of Coldwater Fish in Salt Water 105
115Z:\Customer\CABI\A4337 - Woo\A4419 - Woo Vouchers.vpTuesday, November 05, 2002 11:34:24 AM
Color profile: DisabledComposite Default screen










