1 Structure of the Bacterial Cell - Wiley-VCH · 1 Structure of the Bacterial Cell All bacterial...
28
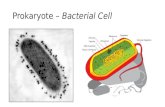
1 Structure of the Bacterial Cell All bacterial cells are surrounded by at least one membrane, the cytoplasmic membrane enclosing the cytoplasm. This simple enclosure can be found only by species living within eukaryotic cells such as Mycoplasma spp. But most cells are surrounded in addition by a thick cell wall (the Gram-positives) and another group by a thin cell wall followed by a second membrane, the outer membrane (the Gram-negatives), where both membranes are separated by the periplasm. Further- more, cells can be surrounded by an extracellular matrix, a capsule or an S-layer, a monomolecular protein layer. In total, Gram-positive bacteria consist of three compartments, the cytoplasm, the cytoplasmic membrane and the extracyto- plasm, while Gram-negatives contain two additional compartments, the peri- plasm and the outer membrane. In addition, cells can contain appendages, flagel- la and/or pili allowing movements and attachment to cell surfaces, respectively. We will present an overview of the composition and function of these different compartments. For details, classic microbiology textbooks should be consulted. 1.1 The Cytoplasm Compartment First of all, the cytoplasm contains all the different types of nucleic acids: the chro- mosome(s) and quite often one or more plasmids and the arsenal of RNAs (mRNA, rRNA, tRNA, sRNA). In addition, the cytoplasm consists of an aqueous solution of salts, sugars, amino acids, vitamins, coenzymes and a wide variety of other soluble materials including about 2000 different protein species. The cyto- plasmic proteins fold into structures that place their hydrophobic segments in the interior to reduce their entropic cost of their exposure to water, and polar residues are generally placed on the protein surface where they can be hydrated. Tradi- tional textbook representations of the prokaryotic cytoplasm show an amorphous amalgamation of proteins in which a randomly arranged chromosome is located. This image of the microbial cell is not correct and, instead, the cytoplasm is highly structured with many proteins carrying out their assigned functions at specific subcellular locations. Therefore, the bacterial cell is more than a bag of proteins. While most proteins may be dispersed throughout the cytoplasm, some seem to 1 Dynamics of the Bacterial Chromosome. Structure and Function. W. Schumann Copyright g 2006 Wiley-VCH Verlag GmbH & Co. KGaA ISBN 3-527-30496-7
Transcript of 1 Structure of the Bacterial Cell - Wiley-VCH · 1 Structure of the Bacterial Cell All bacterial...

1
Structure of the Bacterial Cell
All bacterial cells are surrounded by at least one membrane, the cytoplasmicmembrane enclosing the cytoplasm. This simple enclosure can be found only byspecies living within eukaryotic cells such as Mycoplasma spp. But most cells aresurrounded in addition by a thick cell wall (the Gram-positives) and another groupby a thin cell wall followed by a second membrane, the outer membrane (theGram-negatives), where both membranes are separated by the periplasm. Further-more, cells can be surrounded by an extracellular matrix, a capsule or an S-layer, amonomolecular protein layer. In total, Gram-positive bacteria consist of threecompartments, the cytoplasm, the cytoplasmic membrane and the extracyto-plasm, while Gram-negatives contain two additional compartments, the peri-plasm and the outer membrane. In addition, cells can contain appendages, flagel-la and/or pili allowing movements and attachment to cell surfaces, respectively.We will present an overview of the composition and function of these differentcompartments. For details, classic microbiology textbooks should be consulted.
1.1
The Cytoplasm Compartment
First of all, the cytoplasm contains all the different types of nucleic acids: the chro-mosome(s) and quite often one or more plasmids and the arsenal of RNAs(mRNA, rRNA, tRNA, sRNA). In addition, the cytoplasm consists of an aqueoussolution of salts, sugars, amino acids, vitamins, coenzymes and a wide variety ofother soluble materials including about 2000 different protein species. The cyto-plasmic proteins fold into structures that place their hydrophobic segments in theinterior to reduce their entropic cost of their exposure to water, and polar residuesare generally placed on the protein surface where they can be hydrated. Tradi-tional textbook representations of the prokaryotic cytoplasm show an amorphousamalgamation of proteins in which a randomly arranged chromosome is located.This image of the microbial cell is not correct and, instead, the cytoplasm is highlystructured with many proteins carrying out their assigned functions at specificsubcellular locations. Therefore, the bacterial cell is more than a bag of proteins.While most proteins may be dispersed throughout the cytoplasm, some seem to
1
Dynamics of the Bacterial Chromosome. Structure and Function. W. SchumannCopyright g 2006 Wiley-VCH Verlag GmbH & Co. KGaAISBN 3-527-30496-7

have specific localizations. One example is the B. subtilis replisome which is pre-sent throughout replication of the chromosome within the middle of the cell. Pro-teins can be localized within the cell through a so-called localization tag which isfused either to the N- or to the C-terminus. The most prominent tag is GFP (forgreen fluorescent protein) and its variants. Furthermore, all macromolecular pro-cesses occur in the cytoplasm, such as DNA replication, transcription, translationand recombination.
The Bacterial Cytoskeleton
All eukaryotic cells, from yeast to plants to animals, have an internal frameworkcalled the cytoskeleton. The three types of strut that make up this framework ±microtubules, actin filaments and intermediate filaments ± not only provide cellswith mechanical support, but also serve as tracks for motor molecules to movealong. Bacteria were long thought to lack cytoskeletal filaments, suggesting thatthe cytoskeleton might have evolved after the first primitive eukaryotic cell devel-oped from its bacterial origins. But work carried out over the past few years hasshown that bacteria contain cytoskeletons with homologs to all the eukaryotic cy-toskeletal elements. Bacteria contain a microtubule (tubulin) protein called FtsZ,multiple actin-like proteins including MreB and Mbl, and intermediate filamentproteins such as crescentin in C. crescentus, indicating the very ancient origin ofthese types of intracellular filaments. Strikingly, no nucleators or motor proteinshave been identified for any bacterial cytoskeletal element. In eukaryotes, microtu-bules are involved in various transport processes and, in forming the mitotic spin-dle during chromosome segregation, actin forms a dynamic cytoskeleton involvedin cell shape regulation and mobility as well as the cytokinetic ring during cell di-vision; and intermediate filaments provide mechanical support to the cell and nu-cleus. In procaryotes, the tubulin homolog FtsZ is essential for cell division, form-ing a cytokinetic ring at mid-cell early during the division process (see Section3.3). In association with a number of other cell division proteins, FtsZ then con-stricts concomitantly with septal peptidoglycan synthesis to bisect the cell. Thetwo actin-like proteins, MreB and Mbl, in B. subtilis form helical filaments. Thetwo proteins do not interact to form a hybrid filament but each protein forms a fi-lament of different pitch and length. While MreB forms a short (0.73�0.12 ìm)pitch filament that assembles around the mid-cell position, Mbl filaments (half-life ;8 min) have a longer pitch (1.7�0.28 ìm) and traverse the entire cell length.MreB spirals are dynamic throughout the cell cycle, compacting at the divisionplane in predivisional cells, remaining there until division is complete and thenexpanding to fill the cell. MreB has a role in the maintenance of the cell width andplays a role in chromosome segregation, whereas Mbl is involved in the mainte-nance of the long axis of the cell. Depletion of either MreB or Mbl causes aberrantpartitioning of the origins of replication and a large fraction of anucleate cells.The intermediate filament crescentin (encoded by creS) of C. crescentus is closelyjuxtaposed with the cell membrane and is responsible for the vibrioid shape: celllacking crescentin are rod-shaped. CreS filaments represent true architectural
1 Structure of the Bacterial Cell2

structures within the cell, as their conformation dictates cell shape. And the ParMprotein encoded by plasmid R1 forms F-actin-like cables that actively move plas-mid DNA. In 2001, the three-dimensional structure of MreB of T. maritima wassolved and revealed that it is congruent with that of eukaryotic actins. Later, thestructure of a plasmid-encoded ParM protein was solved and shown to be very si-milar to that of actin and MreB. Both proteins and several others, such as Mbl ofB. subtilis, now belong to the MreB family; and these proteins carry out nucleotidebinding and hydrolysis which regulates polymerization into linear protofilaments.
1.2The Cytoplasmic Membrane Compartment
The cytoplasmic membrane is an 8 nm thick structure enclosing the cytoplasm,separating the interior of the cell from the environment and preventing the diffu-sion of substances into and out of the cytoplasm. It acts as a highly selective bar-rier to concentrate metabolic compounds and nutrients within the cytoplasm andto secrete waste products.
Structure and Composition of the Cytoplasmic Membrane
The cytoplasmic membrane consists of equal amounts of phospholipid and pro-tein. It contains 65±75% of the cellular phospholipids and 6±9% of the cellularprotein. The phospholipids consist of a hydrophobic part (fatty acids) and a hydro-philic part (glycerol), where the glycerol backbone contains two bound fatty acidsand a phosphate group. Three major phospholipid species are present in E. coli,amounting to about 2¥107 molecules per cell: 70±80% phosphatidylethanolamine,15±25% phosphatidylglycerol and 5±10% cardiolipin. All of the phospholipidscontain sn-glycerol-3-phosphate esterified with fatty acids at the sn-1 and sn-2 posi-tions. The predominant fatty acids are the saturated palmitic acid (16:0) and theunsaturated species palmitoleic acid (16:1) and cis-vaccenic acid (18:1). The struc-ture of the membrane is stabilized by hydrogen bridges and hydrophobic interac-tions; and, additionally, cations such as Mg2+ and Ca2+ help to stabilize the mem-brane by forming ionic bonds with the negative charges of the phospholipids.Fatty acid content (chain length, saturation) are dependent on the environmentalconditions, including temperature (see Section 9.3), stage of growth and composi-tion of the growth medium. In particular, membranes have to be maintained in afluid, liquid±crystalline state to allow lateral diffusion of their proteins and proteincomplexes. In addition, fluid membranes have much higher permeabilities tosmall molecules than do gel-phase bilayers. The phospholipids arrange in a lipidbilayer, forming a hydrophobic barrier, preventing the uncontrolled movement ofpolar molecules and allowing the retension of metabolites and proteins.
While some proteins are tightly bound to the membrane with one or moremembrane-spanning domains (so-called integral membrane proteins), others areloosely bound (peripheral membrane proteins) or interact only transiently. Bioin-
1.2 The Cytoplasmic Membrane Compartment 3

formatic analysis of the E. coli proteome indicates that approximately 1000 of the4288 predicted genes encode integral membrane proteins that are critically impor-tant for many cellular functions. However, owing to their hydrophobic and amphi-philic nature, membrane proteins are difficult to study; and they account for lessthan 1% of the known high-resolution protein structures. Topology models de-scribe the number of transmembrane spans and the orientation of the protein re-lative to the lipid bilayer. For E. coli proteins, location of their C-terminus can beeasily determined through the use of the topology reporter proteins alkaline phos-phatase (PhoA) and GFP. PhoA and GFP have opposite activity proliles: PhoA isactive only in the periplasm, whereas GFP is fluorescent only in the cytoplasm.When fused to the C-terminus of a membrane protein, PhoA and GFP accuratelyreport on which side of the membrane the C-terminus is located. The distributionof the membrane proteins changes considerably under different medium andgrowth conditions. The amino acid residues of membrane proteins in the interiorhave similar hydrophobic character, as residues in the interior of the soluble pro-teins and the residues that are exposed to the aqueous environment have the ex-pected polar character. Residues exposed to the nonpolar lipid acyl chains haveeven greater aggregate hydrophobic character than residues in the protein interiorimportant for maintaining the correct conformation and penetration of the pro-tein in the nonpolar lipid bilayer. In general, transmembrane segments are con-sidered to be a-helical. Cytoplasmic membrane proteins can be classified as totheir function they exert:
· proteins involved in energy generation and conservation· proteins involved in solute transport· proteins involved in carbohydrate translocation· proteins and protein complexes involved in the translocation of proteins
through the cytoplasmic membrane· other cytoplasmic membrane proteins.
Proteins Involved in Energy Generation and Conservation
Most biosynthetic and transport processes in eubacteria are driven neither by thehydrolysis of the high-energy phosphate bonds in ATP, GTP or phosphoenolpyru-vate (PEP), nor by coupling to transmembrane ion gradients. Cells growing in fer-mentative conditions (absence of oxygen or other inorganic electron acceptors)produce ATP by substrate-level phosphorylation reactions in the glycolytic path-way. The ATP synthesized by these reactions can be used to form transmembraneion gradients, primarily by reversal of the F1F0 proton-translocating ATPase. Forcells grown in nonfermentive, respiratory conditions, the passage of electronsthrough an electron transfer chain to suitable electron acceptors (oxygen, fuma-rate, nitrate, nitrite, dimethyl sulfoxide [DMSO], trimethylamine N-oxide [TMAO]or hydrogen) is coupled to the extrusion of proteins and the creation of a trans-membrane electrochemical gradient of protons. This proton motive force (PMF,¾ìH) can be used by many transport processes. Bacterial respiratory chains act asa series of physically separate protein complexes. Numerous membrane-bound
1 Structure of the Bacterial Cell4

dehydrogenases transfer two electrons or hydrogen atoms from their specific sub-strates to the pool of quinones, which serve as mobile hydride carriers diffusingthrough the membrane. These quinones shuttle reducing equivalents from thedehydogenases to the terminal reductases or oxidases. While ubiquinone-8 is thepredominant species in aerobic cells, menaquinone-8 is the major species in cellsgrown under anaerobic conditions.
PMF and ATP synthase are involved in energy conservation. The energy re-leased by the passage of reducing equivalents along the electron transfer chains iscaptured in the form of an electrochemical gradient of proteins released upon qui-nol oxidation. PMF consists of two components, an electrical potential owing to theseparation of charge and a chemical gradient or pH gradient, when the external pHdiffers from the internal pH. The magnitude of the electrical potential, ¾é, ismeasured from the distribution of permeant ions, while the pH gradient is mea-sured from the distribution of permeant weak acids or bases. The multisubunitF1F0 proton-translocating ATPase, also called ATP synthase, carries out ATPsynthesis of ATP at the expense of the transmembrane PMF generated in respir-ing cells. Alternatively, ATP synthase can use ATP generated by fermentative sub-strate-level phosphorylation to create the PMF. The enzyme is composed of eightsubunits organized in two distinct complexes, the membrane-embedded F0 com-plex (subunits a, b, c in a 1:2:6±12 stoichiometry) forming a transmembrane pro-ton channel and the peripherally bound F1 complex. The F1 complex consists offive subunits (a3â3�1ä1å1).
Proteins and Protein Complexes Involved in the Translocation of Proteins Throughthe Cytoplasmic Membrane
Integral cytoplasmic proteins and proteins translocated through the cytoplasmicmembrane need assistance by at least one protein. Two major pathways exist totranslocate polypeptide chains through the cytoplasmic membrane, the Sec andthe Tat pathways, accepting unfolded and folded proteins, respectively (see Section8.4), while the insertion of proteins into the cytoplasmic membrane is assisted byYidC alone or in conjunction with the Sec pathway (see Section 8.4.3). In addition,in Gram-negative bacteria, there are protein complexes attached and integratedwithin the cytoplasmic membrane working in conjunction with outer membranecomplexes destined to secrete proteins into the medium (see Section 8.6).
Proteins Involved in Solute Transport
Some small nonpolar and fat-soluble substances, such as fatty acids, alcohols andbenzene, may enter and exit the cell readily by becoming dissolved in the lipidphase of the membrane. Charged molecules, in contrast, such as organic acidsand inorganic salts, which are hydrophilic, do not readily pass the membrane bar-rier but instead must be specifically transported. Water freely penetrates themembrane, since water molecules are sufficiently small and uncharged to passbetween phospholipids. Therefore, polar molecules have to be transported through
1.2 The Cytoplasmic Membrane Compartment 5

the action of membrane transport proteins. Such active transport mechanisms al-low the accumulation of solutes against a concentration gradient. Most solutes aretransported by an energy-dependent process and bacteria produce a remarkablenumber of active transport systems exhibiting high substrate affinity and specifi-city with a broad range of mechanisms of energy-coupling including:
· systems driven by symport or antiport with ion gradients· periplasmic permeases as part of multiprotein complexes that consist of a peri-
plasmic substrate-binding protein and are driven by ATP hydrolysis· ion transport driven by P-type ATPases· serial transport systems that mediate active transport across both the inner and
the outer membrane· group translocation processes that carry out the simultaneous transport and
modification of their substrate.
Serial transport systems can mediate active transport across both the inner and theouter membrane. Since these systems are composed of components residing inthree different compartments (the inner membrane, the outer membrane, theperiplasm), they are explained under Section 1.5. Ion symport systems use a singleintegral membrane protein to couple substrate accumulation to the downhillmovement of a driving ion in the same direction as the substrate. These electro-genic transport processes can be driven by proton or sodium gradients; and thelactose transporter, LacY, is the best studied ion-driven active transport system.The LacY permease contains 12 transmembrane segments of about 20 aminoacids each, with both protein termini facing the cytoplasm. A stoichiometry of oneproton per lactose molecule is seen under most conditions. Other symporters cou-ple substrate accumulation to the sodium ion gradient which is established in en-teric bacteria by the action of sodium/proton antiporters extruding Na+ at the ex-pense of PMF. One example is the MelB protein which transports melibiose andother a-galactosides coupled to Na+ or H+ gradients, while transport of methyl-thio-â-galactoside is coupled only to the Na+ gradient. Antiporters couple the up-take of one compound to the release of a second type. Examples from E. coli arethe potassium/proton exchanger Kha and the two sodium/proton exchangersNhaA and NhaB. Other antiport systems exchange the substrate for a metabolicprocess for its structurally similar product. E. coli cells growing anearobically withfumarate as electron acceptor synthesize a specific C4-dicarboxylate transport sys-tem that exchanges succinate for a molecule of the substrate, fumarate.
ATP-driven transport systems include periplasmic permeases, serial transportsystems and P-type ATPases. Periplasmic permeases exhibiting high affinity fortheir substrates (Km 0.1±1.0 ìM) differ from symporters by virtue of their depen-dence on the high-energy phosphate pool. Their transport activity is relatively un-affected by protonophores dissipating the PMF. Periplasmic permeases, alsocalled ABC (for ATP-binding cassette) transporters appear to be made of four pro-tein domains or subunits: two hydrophobic membrane-spanning domains (MSDs)that are presumed to constitute the translocation pathway or channel across themembrane and two hydrophilic nucleotide-binding domains (NBDs) that interact
1 Structure of the Bacterial Cell6

at the cytoplasmic surface to supply the energy for active transport. ABC transpor-ters now constitute one of the largest superfamilies of proteins known: There are80 ABC transporters in the Gram-negative bacterium E. coli. In the transport clas-sification database (http://www-biology.ucsd.edu/_msaier/transport/), the ABC fa-mily is currently subdivided into 22 subfamilies of prokaryotic importers and 24subfamilies of prokaryotic exporters. All bacterial ABC transporters that mediateuptake utilize a high-affinity solute binding protein which is located in the peri-plasm of Gram-negative bacteria and is either tethered to the cell surface or fusedto the transporter itself in Gram-positive bacteria. These binding protein-depen-dent transporters take up a wide variety of substrates, which include nutrients andosmoprotectants that range from small sugars, amino acids and small peptides tometals, anions, iron chelators (siderophores) and vitamin B12. The best studied ex-ample is the maltose uptake system (Fig. 1.1). Maltose or maltodextrins will bindto the periplasmic maltose-binding protein (MBP, encoded by malE) present in itsopen form. This leads to a conformational change, the closed form, and enables in-teraction with the transporter MalFGK2. Binding to the transporter in turn stimu-lates a conformational change back to the open form, opening the transporter atthe periplasmic site with concomitent release of maltose into the transporter andbinding ATP to the ATP-binding cassette, the MalK protein. Since the open formof MBP has a low affinity towards the transporter, it dissociates, thereby triggeringclosing of the transporter at the periplasmic site and opening at the cytoplasmicsite driven by ATP hydrolysis.
ABC transporters also function in efflux of substances from bacteria, which in-clude surface components of the bacterial cell (such as capsular polysaccharides,lipopolysaccharides, teichoic acid), proteins involved in bacterial pathogenesis(such as hemolysin, heme-binding protein, alkaline protease), peptide antibiotics,heme, drugs and siderophores. E. coli is thought to possess 57 ABC transporters,which can be phylogenetically divided into ten subfamilies. As 44 of these trans-porters have associated periplasmic-binding proteins, they are thought to be in-volved in solute transport.The other 13 transporters are not linked to periplasmic-binding proteins and may therefore be involved in export. CCmABC, which is in-volved in heme export, is the only one for which export function has been identi-fied. The LolCDE complex is a unique lipoprotein-detaching apparatus, but not anexporter. It releases lipoproteins from one leaflet of lipid bilayers but does nottransport them across membranes, representing a new ABC transporter class.
P-type ion-translocating ATPases consist of a large (ca. 100 kDa) protein subunitthat is phosphorylated on an aspartate residue by ATP during the transport cycleand a smaller subunit associated with the catalytic subunit. Examples are multipletransport systems for potassium in E. coli (TrkD, TrkG, TrkH), three magnesiumtransport systems where MgtA and MgtBC mediate only Mg2+ influx while theCorA mediates bidirectional flux of Mg2+ and the uptake of Co2+. A-type ATPasesmediate resistance to toxic oxyanions, such as arsenate, arsenite and antimonite.These plasmid-encoded heavy metal resistance determinants are widespread inbacteria.
1.2 The Cytoplasmic Membrane Compartment 7

Group translocation systems mediate simultaneous transport and modification oftheir substrates. The best characterized systems are PEP:sugar phosphotransferasesystems (PTS), which use a cascade of phosphate transfer reactions from PEP viacommon and sugar-specific proteins to the sugar substrate during transmembraneuptake. Sugar accumulation occurs because the sugar±phosphate product istrapped inside the cell. Furthermore, the PTS plays a key role in catabolite repres-sion and inducer exclusion coordinating and regulating carbohydrate metabolism.
Water fluxes across the cytoplasmic membrane are accomplished by two dis-tinct mechanisms: (1) simple diffusion of water across the lipid bilayer and (2) dif-fusion through water-selective channels called aquaporins (AqpZ). AqpZ mediaterapid and large water fluxes in both directions in response to sudden osmotic up-and downshifts.
1.3
The Cell Wall Compartment
Most bacterial cells are surrounded by a rigid wall that has been thought to deter-mine the shape of the cells (see Section 1.1). In both Gram-negative and -positivecells, the cell wall is located on the outside of the inner membrane, but is furthersurrounded by the outer membrane in Gram-negative bacteria. The major compo-nents of the bacterial cell wall are long glycan strands that are cross-linked byshort peptides containing amino acids in both the d- and l-isoform and the wholeensemble is called peptidoglycan or murein, forming the murein sacculus. The mur-ein of E. coli is composed of glycan strands with alternating N-acetylglucosamineand N-acetylmuramic acid sugar subunits linked together by â1á4 glycosidicbonds. Attached to the carboxyl group of each muramic acid by an amide linkageis a short peptide, l-alanyl-d-isoglutamyl-l-meso-diaminopimelyl-d-alanine, and
1 Structure of the Bacterial Cell8
Fig. 1.1 The maltose uptake system. Maltose(represented by a black dot) is first bound bythe periplasmic maltose-binding protein(MBP) forming a closed complex. Upon con-tact with the maltose transporter composed ofthe integral membrane proteins MalF andMalG, MBP opens to release maltose. Binding
of MBP also triggers a conformational changein the ATP-binding cassette, leading to thebinding of ATP. Next, maltose is transportedinto the cytoplasm, driven by ATP hydrolysis.A.L. Davidson, J. Chen 2004, Annu. Rev. Bio-chem. 73, 241±268; Fig. 1.

the whole unit is called muropeptide. The average glycan strand is bout 30 muro-peptides in length. A molecule of lipoprotein (lpp) is attached to about every tenthmuropeptide; and the covalent link is from the l-carboxyl group of DAP to the ep-silon group of the carboxyl-terminal lysine of the lipoprotein. Further contacts oc-cur with the porins OmpC and OmpF, providing together over 400000 possiblecontacts between the murein sacculus and the outer membrane. Elongation of themurein sacculus is a multisite process, which involves the insertion of strands atabout 200 separate locations simultaneously; and the enzymes responsible movecontinously in one direction around the circumference. The precursors for pepti-doglycan synthesis are a well conserved family of disaccharide pentapeptidescalled lipid II. They are synthesized in the cytosol and then flipped to the outerleaflet of the inner membrane, where they are added to the peptidoglycan mesh-work by two distinct enzyme-catalyzed reactions. One of them, designated trans-glycosylation, adds disaccharide units to extend the linear glycan strand. In theother reaction, transpeptidation, interstrand cross-links form between juxtaposedpeptide side-chains; and this reaction is the target of â-lactam antibiotics. Sincethe transpeptidases were first identified by their ability to bind penicillin, they arefrequently called ,,penicillin-binding proteins,, (PBPs). Most bacterial species codefor multiple PBPs, where E. coli encodes about ten and B. subtilis about 16 PBPs.Peptidoglycan in E. coli is synthesized ubiquitously across the bacterial length,with the exception of the old poles, at which no new peptidoglycan synthesis oc-curs. Gram-positive bacteria contain another major polymer in addition to pepti-doglycan in their cell walls, called teichoic acid. This anionic polymer is essentialfor Gram-positive bacterial cell wall viability, but its precise function is unknown.Bacterial cell walls provide protection against osmotic pressure in the cytoplasmand allow cells to withstand elevated hydrostatic turgor pressure.
1.4
The Outer Membrane Compartment
Gram-negative bacteria are surrounded by an additional membrane layer, the out-er membrane whose most important function is to serve as a selective permeationbarrier. It prevents the entry of noxious compounds and allow the influx of nutri-ent molecules. The outer membrane is an asymmetric lipid bilayer where the in-ner leaflet consists of phospholipids, while the outer leaflet is composed of lipopo-lysaccharide (LPS). As such, lipid bilayers show little permeability for hydrophilicsolutes, including most nutrients. Amino acids, short peptides, sugars or oligosac-charides and other permeants below a threshold size of ~600 Da can cross theouter membrane by diffusion through porin channels. In contrast, other valuableor essential nutrients, including vitamin B12 and iron siderophore complexes, aretoo large and usually too scarce to be aquired effectively through porins. Sub-strate-specific, high-affinity active transporters are used for their uptake located inthe outer membrane. The energy required to allow these transporters to delivertheir substrates into the periplasm derives from a complex of proteins (TonB,
1.4 The Outer Membrane Compartment 9

ExbB, ExbD) in the cytoplasmic membrane (see Section 1.5). Infective agents suchas some colicins and phages evolved to parasitize TonB-dependent systems to killtarget bacteria.
The outer membrane of Gram-negative bacteria such as E. coli contains threemajor classes of proteins: lipoproteins, â-barrel proteins and multicomponent sur-face structures, such as pili and flagella. Liporoteins have lipid covalently attachedto the N-terminal cysteine that anchors these molecules in the inner leaflet of theouter membrane. The â-barrel proteins are composed of â-sheets that are wrappedinto cylinders (Fig. 1.2). Because of this structure, many of these proteins, alsocalled porins, form channels allowing the influx of nutrients and the extrusion ofwaste products. These porins can be classified in two groups: nonspecific and spe-cific channel-forming proteins. Nonspecific porins such as OmpF act as molecu-lar sieves, while the others, like LamB, exhibit substrate specificity, in this case formaltose polymers. Since the â-barrel proteins are exposed on the surface, theymay be used as phage receptors, such as OmpF for K20 and LamB for �. The thirdmajor class of outer membrane proteins, the pili, serve to attach bacteria to sur-faces. Each of the many different types of pili has specific affinity for certain sur-faces (see below).
Composition and Biological Function of Lipopolysaccharide
Lipopolysaccharide (LPS) is a unique constituent of the bacterial outer membraneand is composed of three components: the proximal, hydrophobic lipid A region,the distal, hydrophilc O-antigen polysaccharide region that protrudes into themedium, and the core oligosaccharide region that connects the other two. Lipid Ais a polar lipid of unusual structure, in which a backbone of glucosaminyl-â-(1á6)-glucosamine is substituted with six or seven fatty acid residues, all of them
1 Structure of the Bacterial Cell10
Fig. 1.2 Structure of the OmpF porin. View of the monomer from the side(left) and view of the trimer from the top. The loop inside the opening narrowsthe channel. H. Nikaido 2003, Microbiol. Mol. Biol. Rev. 67, 593±556; Fig. 2.(This figure also appears with the color plates.)

saturated. LPS biosynthesis begins in the bacterial cytoplasm as the acylation ofuridyl-diphospho-N-acetylglucosamine (UDP-GlcNAc) with â-hydroxymyristate.After deacetylation, the product of this reaction is further modified with â-hydro-xymyristate to generate UDP-2,3-diacylglucosamine. Cleavage of the pyrophos-phate bond produces 2,3-diacylglucosamine-1 phosphate and, after condensationof this compound with another molecule of UDP-2,3-diacylglucosamine and 4¢phosphorylation, the intermediate lipid IVA is formed. Two Kdo (3-deoxy-d-man-no-oct-2-ulosonic acid) residues are then transferred to lipid IVA. Two acetyltrans-ferases, HtrB and MsbB, add lauroyl and myristoyl groups. The core sugar resi-dues are added on this intermediate and the MsbA ABC transporter is thought totranslocate these molecules from the cytoplasmic side to the periplasmic surfaceof the plasma membrane. The O-antigen is added after the MsbA-mediated trans-location. Subsequent transport reactions move the LPS molecules across the peri-plasmic space into the inner leaflet and finally to the outer leaflet. Lipid A is thebiologically active component of LPS which causes inflammation and septic shockthrough Toll-like receptor 4 recognition.
Three kinds of LPS modifications have been observed: (1) substitution of thephosphate group at the 4¢ position with phosphoethanolamine providing resis-tance against polymyxin B, a lipid A-binding cationic cyclic peptide antibiotic, (2)l-Ara4N decoration which is induced by activation of PrmA forming a two-compo-nent system (see Section 6.2.9) with its cognate histidine kinase, PrmA, and (3)addition of palmitate via acyloxyacyl linkage at position 2. LPS palmitoylation pro-vides resistance against cationic antimicrobial peptides induced by the innate im-mune response to bacterial infections. These modifications may occur alone or incombination on a single LPS molecule, yielding at least six different LPS species.
Gram-negative bacteria have long been known to produce membrane blebs orvesicles of 0.5±1.0 ìm into the culture medium when bacteria where grown in ly-sine- or phosphate-limited conditions or when protein synthesis was inhibited.The membrane component of these vesicles is derived from the outer membrane.The naturally competent Neisseria and Haemophilus spp produce large amounts ofvesicles in culture to transfer DNA. Commensal Bacteroides spp of rumen intes-tine produce vesicles containing cellulase and xylanase which can provide the cellswith a source of carbon from the digestion of nonmetabolizable carbohydratepolymers. Very recently, it has been shown that bacterial vesicles have an impor-tant biological function in the secretion and delivery of bacterial protein toxins tomammalian cells (see Section 8.6.6).
Outer Membrane Proteins
Outer membrane proteins (OMPs) are synthesized in the bacterial cytoplasm asprecursors with N-terminal signal peptides and then translocated across the cyto-plasmic membrane via the Sec pathway (see Section 8.4.1). After removal of thesignal peptide by the signal peptidase, the mature proteins assume â-barrel struc-tures with hydrophobic outer surfaces, suitable for interaction with LPS or othermembrane lipids. Several factors are involved in the folding of OMPs, where most
1.4 The Outer Membrane Compartment 11

of these are only in part required for the targeting of OMPs and presumably fulfilloverlapping or partially redundant functions. Outer membrane proteins can beclassified into several classes depending on their function:
· lipoproteins· general porins· substrate-specific porins· other outer membrane proteins.
Lipoproteins
More than a dozen lipoproteins have been described where the murein lipopro-tein (gene lpp) is the most prominent and best studied member. Lpp is a smallprotein (7200 Da) present in approximately 7¥105 copies per cell. Its N-terminalcysteine is modified at two sites. Its sulfhydryl group is substituted with a digly-ceride and its amino group by a fatty acyl residue, allowing penetration into the in-ner leaflet of the outer membrane. About one-third of the murein lipopotein mo-lecules are bound covalently to the peptidoglycan layer through the å-amino groupof its C-terminal lysine, thereby fixing the outer membrane to the murein saccu-lus. Deletion or mutations of lpp result in numerous defects such as periplasmicleakage, increased susceptibility to many toxic compounds and the formation ofmembrane vesicles.
General Porins
The general diffusion pores formed by porins allow the diffusion of hydrophilicmolecules (<600 Da) and exhibit no particular substrate specificity, despite someselectivity for either anions or cations. They form water-filled channels across theouter membrane and are either homotrimers formed by three hollow â-barrels,such as OmpC, OmpF and PhoE, or monomeric, including OmpA and OmpG.The classic porins OmpC, OmpF and PhoE have 18 antiparallel â-strands each.The OmpC and OmpF porins are among the most abundant outer membraneproteins and their expression is extensively regulated. OmpC, with a pore dia-meter of 1.08 nm, is thought to be important in environments where nutrientsand toxin concentrations are high, such as in the intestine, and it is the predomi-nant porin at high temperatures and high osmolarities. The three-dimensionalstructure of OmpF has been determined and the most conspicuous structural fea-ture is the presence of an ,eyelet, region, a narrow site constricting the pore andlined with charged residues. These residues are expected to cause a strong trans-versal electric field, in addition to a screw-like field in the wider parts of the pore.This feature is conserved in general diffusion porins.
OmpF, which has a larger pore diameter (1.12 nm) which results in a 10-foldfaster diffusion rate, is thought to be important in habitats where nutrient and tox-in concentrations are low, such as in fresh water, and it is more abundant at lowtemperatures and low osmolarities. Both OmpF and OmpC play an important rolein maintaining the periplasm and cytoplasm in an iso-osmotic state. Both proteins
1 Structure of the Bacterial Cell12

are present at 105 molecules per cell. While in low-osmolarity medium OmpF pre-dominates, OmpC is present at enhanced levels in high-osmolarity medium at theexpense of OmpF; and the relative amount of each protein in response to the ac-tual medium osmolarity is regulated by the EnvZ±OmpR two-component signaltransduction system (see Section 6.2.8.3).
The trimeric phosphoporin PhoE is produced only under conditions of phos-phate starvation. The channel-forming motif of PhoE is a 16-strand anti-parallel â-barrel. Short â-hairpin turns define the periplasmic end of the barrel, whereaslong irregular loops are found at the cell surface.
OmpA (35 kDa) inserts as a monomer into the outer membrane and is presentin about 100000 copies per cell. OmpA contains a nonspecific diffusion channelwith a pore diameter of 0.7 nm; and the penetration of solutes through the OmpAchannel is about two orders of magnitude slower than that through the OmpFchannel. OmpA is composed of two domains, an N-terminal membrane-em-bedded domain of 170 amino acid residues, serving as a membrane anchor, and aC-terminal 155-residue domain located in the periplasmic space and proposed tospecifically interact with the peptidoglycan layer. The N-terminal domain consistsof eight antiparallel â-strands that are connected by three short periplasmic turnsand four relatively long surface-exposed loops forming a hydrophobic protein sur-face and a polar interior, where the barrel interior accomodates several smallwater-filled cavities. But no continous transmembrane channel could be detectedquestioning the relevance of OmpA in pore formation. Mutants lacking OmpAare extremely poor recipients in conjugation experiments and they tend to pro-duce spherical cells.
The monomeric OmpG is able to rescue the growth of porin-deficient bacteriaon media containing maltodextrins as large as maltopentose as the sole carbonsource. The physiological role of OmpG is not clear. OmpG is not detected in theouter membrane of several E. coli strains, such as K-12, by Western blot analysis.But the ompG gene is detected by PCR analysis in the genome of these strains, in-dicating that the lack of expression may be due to the growth conditions.
Substrate-specific Porins
Besides general diffusion pores that discriminate between solutes (if at all) ontheir charge and size, there is a second class of porins which recognize their sub-strate. Among them are LamB of E. coli and ScrY of Salmonella typhimurium. ThelamB gene is part of the mal regulon (see Section 6.2.8.2) which is induced by mal-tose or maltodextrins. The LamB protein forms homotrimers whose monomersconsist of 18-stranded antiparallel â-barrels. The substrate translocation pathwayinvolves a row of aromatic amino acids (greasy slide) that is lined up by polar resi-dues (ionic track). Sugar residues (maltose or maltodextrins up to maltoheptaose)are in van der Waals, contact with the greasy slide by their hydrophobic face, whilehydrogen bonds are formed between their hydroxyl groups and the ionic track re-sidues. Movement of the sugar through the channel proceeds by continous dis-ruption and formation of these hydrogen bonds. In a chemostat study under car-
1.4 The Outer Membrane Compartment 13

bohydrate-limiting conditions, the LamB production became strongly derepressedand conferred growth advantages at limiting concentrations of not only glucosebut also lactose, arabinose and even glycerol (in addition to maltose, trehalose,melibiose). In summary, the LamB channel is not really a maltose-specific chan-nel, but it facilitates the influx of a wide variety of carbohydrates when they existin low concentrations in the environment. LamB serves also as the receptor forbacteriophage �. Other specific channels are ScrY (allows the rapid diffusion of alarge varieties of sugars, such as glucose, fructose, arabinose, maltose, lactose, raf-finose, sucrose, maltodextrins), BglH (aryl-â-d-glucosides), Tsx (nucleoside trans-port) and FadL (utilization of long-chain fatty acids as a carbon source).
Other Outer Membrane Proteins
Since the outer membrane acts as a major barrier when proteins have to be ex-ported, we need specific mechanisms to overcome this barrier. Currently, at leastfive different mechanisms are known to be utilized and these are described in de-tail under Section 8.6. Export channels of the type I machinery make use of theouter membrane channel TolC, while those of type II are called secretins. Compo-nents of P and type I pili are secreted across the outer membrane, using an usherwhich forms ring-like structures, 15 nm across, and contains a central pore.
OmpT of E. coli is a surface membrane protease and is the prototypical memberof the omptin family of Gram-negative bacteria. OmpT catalyzes the activation ofplasminogen to plasmin, a function that is physiologically relevant for the viru-lence of Y. pestis and for clinical E. coli isolates. OmpT also plays a role in bacterialvirulence by the cleavage of protamine and other cationic peptides with antibioticactivity. The protease folds into a 10-strand antiparallel â-barrel conformation withextracellular loops that extend well beyond the membrane. The active site is lo-cated within a deep groove formed by loops L4 and L5 on the one side and L1, L2,and L3 on the other. The structure also revealed a binding site for a single LPSmolecule that appears to be important for the catalytic activity of the enzyme.OmpT seems to function through a novel mechanism involving an Asp210±His212 catalytic dyad that, together with Asp83±Asp85, activates a putative nucleo-philic water molecule. OmpT cleaves substrates between dibasic residues withhigh catalytic efficiency, which has been shown to be important for the inactiva-tion of antibiotic peptides and colicins and the degradation of some recombinantproteins expressed in E. coli. It copurifies with protein inclusion bodies and re-tains activity under denaturating conditions, including boiling or in the presenceof up to 4 M urea; and therefore it can be a major source of protein degradationduring the solubilization and renaturation of inclusion bodies.
What is known about Omp biogenesis? Omps fold in the periplasm before theirinsertion into the outer membrane. In vitro, LPS stimulates the folding of Omps;and the periplasmic chaperone SurA also stimulates Omp folding, while anotherperiplasmic protein, Skp, plays an as yet unidentified role. The insertion of pro-teins into membranes generally requires a proteinaceous machinery, but no com-ponents of such a putative machinery have been identified. A protein possibly ful-
1 Structure of the Bacterial Cell14

filling these criteria is the surface antigen designated Omp85 in Neisseria spp andD15 in Haemophilus spp. Genes coding for Omp85/D15 homologs are present inall Gram-negative bacteria examined. The omp85 gene is essential and, uponOmp85 depletion, unassembled forms of various outer membrane proteins accu-mulate within the periplasm.
What is known about the dynamics of the outer membrane surface in live cells?To answer this question, the spatial distribution of LamB found mainly as a tri-mer in the outer membrane (there are about 30000 monomeric copies) has beenstudied using fluorescently labeled phage � tails. It turned out that there are twopopulations, mobile (~40%) and immobile LamB trimers. The mobile LamB tri-mers move along the bacterial surface and these movements are restricted by anunderlying dynamic spiral pattern. The distribution of the protein changes withintens of seconds.
R. Koebnik, et al. 2000 Structure and function of bacterial outer membrane proteins: barrels ina nutshell. Mol. Microbiol. 37, 239±253.
1.5
The Periplasmic Compartment
The periplasmic space extends between the inner and outer membranes of Gram-negative bacteria. The architecture of this compartment facilitates cell wall andouter membrane growth and coordinates these processes with cell division. Pro-teins residing in the periplasm fulfil important functions in the detection and pro-cessing of essential nutrients and their transport into the cytoplasm. Furthermore,periplasmic proteins promote the biogenesis of proteins entering this compart-ment along with components destined for incorporation into the peptidoglycan,outer membrane or capsules. In addition, the periplasm can contain glucanswhich play an important role in adaptation to hypoosmotic conditions.
The width of the periplasmic space has been determined to vary over 13±25 nm, depending on the growth conditions. Located within this region is thepeptidoglycan layer, with an estimated thickness of 5±8 nm, which is in close as-sociation with the outer membrane. Attachment of these two structures is facili-tated by major outer membrane lipoprotein, Lpp, and by strong interaction of theporins with the peptidoglycan.
Periplasmic Glucans
Periplasmic glucans, also called MDOs (membrane-derived oligonucleotides), areimportant components of the bacterial envelope of many Proteobacteria. Glucoseis the sole sugar component and residues are linked essentially by â-glycosidicbonds. Glucans of E. coli are heterogeneous in size, varying from five to 12 glu-cose residues, with the principal species containing eight to nine glucose residues.Their structures are highly branched, the backbone consisting of â-1,2 linked glu-
1.5 The Periplasmic Compartment 15

cose units to which the branches are attached by â-1,6 linkages. These polymersare substituted to various degrees by sn-1 phosphoglycerol, phosphoethanolamine,phosphoethanolamine and succinic acid O-ester. In E. coli, the opgGH operon en-codes two proteins (OpgG, OpgH) required for the assembly of the glucan struc-ture. OpgH, a glycosyl-transferase anchored in the cytoplasmic membrane cata-lyzes linear oligomers of â-1,2-linked glucose units synthesized from UDP-glu-cose. OpgH could form a channel for glucan translocation from the cytoplasm tothe periplasm during synthesis. OpgG is a periplasmic protein whose functionmight be to catalyze the addition of branches to the linear backbone emerging onthe periplasmic side of the inner membrane. The two inner membrane proteinsOpgB and OpgC have been implicated in glucan substitutions. While OpgB is aphosphoglycerol transferase with its activity located at the periplasmic face of thecytoplasmic membrane, OpgC is required for the periplasmic succinylation of theglucans. The opgGH operon is under osmotic control and many Gram-negativebacteria respond to hypoosmotic conditions by synthesizing large amounts ofperiplasmic glucans. Under conditions of low osmolarity they can account for upto 5% of the dry cell weight. The nature of the osmosensor is currently unknown.
Periplasmic Proteins
Periplasmic proteins can be divided into several categories based on their functions:
· solute or ion-binding proteins that function together with ABC transporters orchemotaxis receptors for the sensing and uptake of amino acids, vitamins, su-gars, peptides and ions;
· catabolic enzymes that degrade complex molecules into smaller ones for trans-port through the inner membrane;
· detoxifying enzymes protecting cells from toxic compounds;· enzymes involved in the biogenesis of envelope proteins and proteinaceous ap-
pendages, LPS, peptidoglycan, capsules and MDOs;· serial transport systems.
Periplasmic binding proteins concentrate the solutes due to their high affinity fortheir ligand (in the range 0.1±1.0 ìM) and their high concentration (up to1.0 ìM). Binding proteins are monomers generally consisting of two globular do-mains with a molecular mass of 20±40 kDa. Crystal structures of several bindingproteins revealed that they are ellipsoidal and consist of two lobes connected by aflexible linker. These lobes are apart in the unliganded state and they cometogether to bind the solute, using hydrogen bonds. The complex binding protein±ligand interacts with the cognate membrane permease to promote the release ofsolute and its transport across the inner membrane. Four binding proteins play arole in chemotaxis (see Section 6.2.9): MalE interacts with Tar (specific for serine),MglB and RbsB with Trg (ribose and galactose) and DppA with Tap (peptides).
Polypeptides emerging on the outer surface of the inner membrane must foldinto their native state and assemble into quaternary structures in the case of oligo-meric proteins. Proteins destined for the outer membrane may be either released
1 Structure of the Bacterial Cell16

as periplasmic intermediates before insertion or may be translocated directly fromthe inner to the outer membrane. Correct folding of periplasmic and outer mem-brane proteins is aided by protein disulfide isomerases catalyzing disulfide bondformation and proline isomerases (see Section 7.3). Some of these proteins mayalso act as molecular chaperones.
Serial Transport Systems
While most nutrients gain access to their transporters in the cytoplasmic mem-brane by diffusion through porins across the outer membrane (see above), a fewsubstrates are too large to effectively enter by this route. Examples are the ferricsiderophore complexes and vitamin B12. Two serial transport systems, also calledgated channels, have been described as being involved in the uptake of these com-pounds: TonB/ExbB/ExbD and Tol/Pal, which energize the transport via thePMF. Gram-negative bacteria contain several high-molecular-weight outer mem-brane proteins, so-called TonB-dependent receptors. Examples of such receptorsinclude BtuB for vitamin B12 and six receptors for Fe3+-siderophore complexes, in-cluding FhuA, FecA and FepA. They are involved in the uptake of large substrates,including iron-siderophore complexes (iron chelators of microbial origin) and vita-min B12. These receptors bind to the ligands with high affinity and their functionrequires an interaction with the periplasmic protein TonB, which either spans thethickness of the periplasm or shuttles between the inner and outer membrane.The action of TonB requires that the cytoplasmic membrane is energized and thatthe energy is transferred to the receptors with the assistance of two cytoplasmicmembrane proteins, ExbB and ExbD. The cellular ratio of the three proteins Ton-B:ExbB:ExbD is 1:2:7; and this stoichiomeric information predicts a complex of~260 kDa.
How do these receptor proteins work? When the crystal structures of severalsiderophore receptors and BtuB were solved, the â-barrel in all these monomericproteins consists of 22 strands and the N-terminal portion, consisting of about150±200 residues, was found as a globular domain that is inserted into the barrelfrom the periplasmic side, forming a plug. Binding of a ligand induces a confor-mational change within the plug, so that the most N-terminal portion containinga short motif, the so-called TonB box (interacts with TonB) can interact withTonB. This first step does not create a large channel, but is followed by a large-scale conformational change caused by the energized TonB.
Two major models have been proposed for how TonB might act as a energytransducer: the propeller and the shuttle models. In the propeller model, TonB re-mains associated with the complex at all times and undergoes a rotary motion ofthe C-terminal propeller, initiated by ExbB, ExbD and the PMF. When the propel-ler becomes associated with the barrel of the outer membrane transporter, the ro-tary motion initiated at the inner membrane causes the outer membrane trans-porter to release its bound ligand into the periplasmic space. In the shuttle model(Fig. 1.3), TonB starts in an unenergized conformation in complex with ExbB andExbD. ExbBD use the PMF to convert TonB to what is believed to be an energized
1.5 The Periplasmic Compartment 17

conformation. Then, TonB diffuses to the outer membrane where it docks toeither OmpA or Lpp waiting for transporters to signal ligand occupancy. Aminoacid 160 of TonB interacts with the Ton box of the outer membrane transporter.Subsequently, TonB is recharged.
The Tol±Pal system forms a trans-envelope bridge linking inner and outermembranes and the peptidoglycan layer and is composed of seven proteins(Fig. 1.4). TolQ, TolR and TolA are inner membrane proteins that interact to-gether by their transmembrane helices. The periplasmic TolB and the lipoproteinPal form another complex anchored to the outer membrane. TolB interacts in aPal-dependent manner with OmpA and Lpp, while Pal forms a homodimer in thecell envelope and interacts with OmpA. Pal is a lipoprotein with a serine residueat position +2 that results in its localization in the outer membrane. After cleavageof the signal sequence by the signal peptidase II and acylation, Pal is targeted tothe outer membrane by the LolABCDE system (see Section 8.5). The peptidogly-can-binding sequence of Pal is located between residues 97 and 114 and forms aproposed a-helical motif.
Two functions have been suggested for the Tol±Pal apparatus. First, this systemmay contribute to maintaining cell envelope integrity through the interactions be-tween the TolB and Pal proteins with the murein layer and the OmpA and Lppproteins. Second, the Tol±Pal system might play a more dynamic function in cell
1 Structure of the Bacterial Cell18
Fig. 1.3 Shuttle model for TonB-dependentenergy transduction. Uncharged TonB (1) isenergized by uptake of a proton (2) and shutt-les to the outer membrane where is docks toOmpA/Lpp waiting for a transporter such asFepA with a bound ligand. Upon interactionwith the Ton box of the the transporter the
conformational energy is transduced to thetransporter (3) triggering uptake of the ligandinto the periplasm (4). The discharged TonBshuttles back to the ExbBD complex to be-come recharged. K. Postle, R.J. Kadner 2003,Mol. Microbiol. 49, 869±882; Fig. 2. (This figu-re also appears with the color plates.)

envelope biogenesis. It might function to bring the inner and outer membranesin close proximity via the TolA C±terminal domain±Pal interaction. The centraldomain of TolA, which forms an extended amphipatic a-helix, could be requiredfor the transport of outer membrane components through the aqueous periplasmto reach the inner leaflet of the outer membrane. But it remains to be analyzedwhether the PMF-dependent energization of the Tol±Pal system is directly in-volved in maintaining the cell wall architecture.
1.6
Extracellular Matrices
Surface Layers
Surface layers (S-layers) have been found in up to 400 different species of botheubacteria and archaea as the outmost cell envelope layer. They consist of onespecies of (glyco)protein, the S-protein, which assembles into characteristic two-dimensional crystalline layers (lattices) at the cell surface. This assembly is an en-tropy-driven process during which individual S-protein monomers form multipleinteractions with each other and with the underlying cell envelope. The latticescan be quite porous, with pores occupying up to ~70% of their surface. The poreswithin an S-layer are of identical size (usually in the 2±8 nm range) and shape;and two or more distinct classes of pores can be present. S-layer proteins areamong the most abundant cellular proteins and the S-layers fulfill many roles forthe cell. They function as protective coats, as molecular sieves, as structures in-volved in cell adhesion and surface recognition, as scaffolding for enzymes, as
1.6 Extracellular Matrices 19
Fig. 1.4 The Tol±Pal serial transport system. The topologies of the differentprotein components are shown and interactions between these componentsindicated by arrows. E. Cascales, R. Lloubes 2004, Mol. Microbiol. 51, 873±888; Fig. 9.

virulence factors and as molecular and ion traps. Furthermore, they act as viru-lence factors in pathogens, mediate resistance against bactericidal complement ac-tivity and adhesion to extracellular matrix proteins and temper the proinflamma-tory cytokine response. In summary, S-layers presumably evolved by selection inresponse to specific environmental and ecological pressures.
The molecular mass of S-layer proteins ranges from 40 kDa to 170 kDa, andsome of them can become glycosylated (see Section 6.5.3). These proteins are of-ten weakly acidic, typically contain 40±60% hydrophobic amino acids and possessfew or no sulfur-containing amino acids. The S-proteins from Gram-negative bac-teria interact with the sugar moiety of the LPS component of the outer mem-brane. It has been determined that S-proteins constitute between 5±10% of the to-tal protein content of the cell, and ~5¥105 molecules are required to surround anaverage-sized bacterial cell. Most S-layer genes are monocistronic units expressedfrom one or more strong promoters. In most cases, the mRNA contains an un-translated 5¢ region (ranging over 33±358 bp) which can fold into stable secondarystructures causing a half-life of the S-layer mRNA from 10 min to 22 min. Thebiosynthesis of S-layers is a very complex process in which the amount of the pro-tein component, its translocation through the cell wall and its incorporation intothe existing S-layer lattice have to be coordinated with the growth rate of the bac-terium and the synthesis of other cell wall components. To keep the surface of abacterium with an average generation time of 20 min completely covered with theclosed protein lattice, approximately 500 S-layer subunits have to be synthesizedper second. Some eubacterial species can produce two superimposed S-layers;and each is usually composed of a different subunit species.
U.B. Sleytr, T.J. Beveridge 1999, Bacterial S-layers, Trends Microbiol. 7, 253.
Capsules
Many eubacteria secrete on their surfaces slimy materials which in most casesconsist of polysaccharides and, in a few cases, protein. These layers are called cap-sules and vary in different microorganisms; but they usually contains glycopro-teins and a large number of different polysaccharides, including polyalcohols andamino sugars. Capsule layers have several functions:
· They play an important role in the attachment of certain pathogenic organ-isms to their hosts.
· They provide protection against phagocytic cells and the immune system.The most prominent case is that of S. pneumoniae where capsulated cells in-jected into a mouse will cause death in a few days.
· Since they bind a significant amount of water, they play some role in resis-tance to desiccation.
Capsular polysaccharides are high-molecular-weight acidic polymers composed ofoligosaccharide repeating subunits and are found on bacterial cell surfaces andclassified into four groups. In E. coli, the group-1 K capsular serotype of each
1 Structure of the Bacterial Cell20

strain is dependent upon its repeat unit structure. Approximately 80 different Kantigens have been identified in E. coli, reflecting differences in the structure andimmunochemistry of the capsular polysaccharide repeat units. Here, the E. coliK30 serves as the prototype. K-antigen biosynthesis involves the production of un-decaprenol pyrophosphate-linked repeat units by glycosyltransferases acting at thecytoplasmic face of the inner membrane. The repeats units are thought to be ex-ported across the inner membrane involving a putative flippase (Wxz), where theyare polymerized by a reaction requiring the wzy gene product. The majority of theundecaprenol pyrophosphate-linked repeat units are used for polymerization ofcapsular polysaccharide and require a tyrosine autokinase, Wzc, and its cognatephosphatase, Wzb. After polymerization, the capsule is transported to the cell sur-face in a process that involves the outer membrane lipoprotein Wza. This lipopro-tein forms multimeric structures resembling secretions for type II and type IIIprotein secretion (see Sections 8.6.2, 8.6.3) acting as channels in the outer mem-brane to allow the polymers to reach the cell surface. It is not yet clear how thepolymer is moved through the periplasm to these channels, nor how it is as-sembled on the cell surface to form the capsular structure. Group 1 capsules af-ford protection from the host immune system by impeding phagocytosis. Theyare also involved in an intimate association with the glycocalyx of epithelial cellsand probably contribute to biofilm development, but initial attachment to theepithelium also requires the participation of pili.
1.7
Appendages
Various types of exocellular appendages are important accessory components forbacteria to best fit their particular ecological niche. These appendages consist of fla-gella and pili (also called fimbriae). While flagella are involved in bacterial motility,pili mediate adhesion to specific targets in the environment. Both flagella and piliare anchored through the cell body to both periplasmic and membrane components.
Flagella
Flagella (singular: flagellum) are long, thin (about 20 nm diam.) appendages freeat one end and attached to the cell at the other. They consist of the three maincomponents basal body, hook and filament and function in the movement andchemotaxis of bacterial cells (Fig. 1.5). While some bacterial species contain onesingle, polar flagellum, others contain multiple flagella at one pole and othersmany flagella around the whole cell (peritrichous). Flagella are able to rotate ex-erted by a proton- or sodium-driven rotary motor that switches between counter-clockwise and clockwise rotation.
The bacterial flagellum is composed of over 20 protein species with approxi-mately another 30 proteins required for regulation and assembly; and it repre-sents the most complex of all prokaryotic organelles (Fig. 1.5). The filament is ty-
1.7 Appendages 21

pically about 20 nm in diameter and usually consists of thousands of copies of asingle protein called flagellin. In some cases, the filament is composed of severaldifferent flagellins. At the tip of the flagellum is a capping protein termed HAP2,forming a pentamer. New flagellin subunits assemble at the end of the filamentunder the cap protein, forming a plate-like structure with five legs protrudingdownwards and interacting with the filament. As each subunit is added, the caprotates along the end of the helical filament, with a complete rotation of the capoccuring for every 55 flagellin subunits added. The filament is connected to thebasal body by the hook, composed of a single protein, FlgE. The hook has a de-fined length of 55 nm and consists of about 120 subunits of FlgE. Hook length iscontrolled by the C (cytoplasmic) ring located beneath the basal body and com-posed of the three switch proteins. The C ring is filled with FlgE subunits, actingin the capacity of a measuring cup and then exported and assembled. Therefore,along the model, the length of the hook is determined by the capacity of the Cring. The two hook-associated proteins HAP1 and HAP3 are located at the hook±filament junction. The basal body consists of a rod, several rings, the Mot pro-teins, the switch complex and the flagellum-specific export apparatus. The ringsanchor the flagellum to the cytoplasmic membrane (MS ring), the peptidoglycan(P ring) and the outer membrane (L ring) in Gram-negative bacteria. The flagellaof Gram-positive bacteria are anchored only through the MS ring in the cytoplas-mic membrane. The three switch proteins FliG, FliM and FliN allow the flagel-lum to switch rotation from clockwise to counterclockwise. This allows E. coli cellsto chance direction of swimming in response to attractants or repellents sensed
1 Structure of the Bacterial Cell22
Fig. 1.5 Composition of the bacterialflagellum. The flagellum consists of thethree components basal body (anchorsthe organelle within the cell envelopeand functions as the switch), the hookand the filament, acting as a propeller.S.L. Bardy, et al. 2003, Microbiology 149,295±304; Fig. 1.

by a complex chemotaxis system where the phosphorylated CheY protein contactsFliM (see Section 6.2.9). Assembly of the whole flagellum occurs through the typeIII secretion pathway (see Section 8.6.3).
Salmonella cells are approximately 0.73 ìm wide and 1.4 ìm long. Each cell pos-sesses about five flagella and these flagella are arranged peritrichously (lateral)around the cell. Each cell rotates its flagella at ~200 revolutions s±1 and swims atspeeds of ~55 ìm s±1. This is a propulsion rate of many cell body lengths per sec-ond. E. coli cells are similar: ~270 revolutions s±1 and 36 ìm s±1. In comparison,the marine bacterium V. alginolyticus, whose cell size is similar to the cell size ofE. coli and Salmonella, swims at speeds as fast as 116 ìm s±1 and its flagellar rota-tion rate is ~1000 revolutions s±1.
High viscosity generally impedes flagellar rotations. As a consequence, bacteriaimplement additional strategies to maximize movement in viscous conditions andon surfaces, including the number of flagella and their arrangement. Many peri-trichous bacteria upregulate the number of flagella and alter extracellular compo-nents such as polysaccharide and surfactant production to enable movement onsurfaces, called swarming. Other bacteria induce completely new, alternative fla-gellar systems in response to growth in viscous environments and on surfaces.They are polarly flagellated when they grow in liquid medium and have mixed(polar and peritrichous) flagella when they are grown on surfaces. One example isthe above mentioned V. alginolyticus, where the peritrichous flagella enable effec-tive motility in highly viscous environments (20 ìm s±1 in a ~200-cp environ-ment). B. subtilis possesses two types of flagellar motors driven by different energysources (the sodium and proton motive forces) but only one set of flagellar genes.
Pili (Fimbriae)
Pili (singular: pilus), also called fimbriae, are hair-like appendages built by proteinsubunits called pilin or fimbrin and usually extend 1±2 ìm from the surface ofGram-negative bacteria, with a diameter ranging from 2 nm to 8 nm. They func-tion in bacterial cell-to-cell interactions, adhesion to specific receptors of hostcells, either uptake or transfer of genetic material and twitching motility, a formof locomotion that is powered by extension and retraction of the pilus filament,and they provide receptors for bacteriophages. Pili can be classified on the basis oftheir physical properties, antigenic determinants or adhesion characteristics.Class I and IV pili represent the two best characterized model systems and areubiquitously present among Gram-negative pathogens. Pili are assembled by sev-eral distinct pathways and we will explain the biogenesis of F-pili, P-pili, type 4pili, curli and T-pili.
F-pilus Biogenesis
The F-pilus with a length of about 1 ìm is encoded by the tra genes located on the100-kb F factor. It has a specialized tip structure, the nature of which remains un-known, and a helical array of subunits of 8 nm diameter with a 2-nm lumen. The
1.7 Appendages 23

tip is involved in the recognition of a suitable recipient cell as well as F-specific fi-lamentous phages such as M13, f1 and fd. RNA phages including R17 and Qâ at-tach to the sides of the pilus. The F-pilus is composed of F-pilin encoded by traAand translated as a 121-amino-acid propilin polypeptide. Correct insertion of thepropilin into the membrane requires TraQ, an inner membrane protein. Propilinis processed by the signal peptidase (lepB) to a 77-amino-acid intermediate that isacetylated at its N-terminus by TraX. The mature F-pilin accumulates in the innermembrane as a pool of ~100 000 subunits. The F-pilus is assembled from thispool by eleven tra gene products (TraL, -E, -K, -B, -V, -C, -W, -F, -H, -G, TrbC).While TraA, -L, -E, -K, -D and -G are involved in assembling a structure at the cellsurface, thought to be the pilus tip, TraB, -V, -W, -F and -H are involved in pilusextension, which is stabilized by TraP. TraU appears to be involved in DNA trans-fer and TrbI in pilus retraction after contact with the recipient cell.
P-pilus Biogenesis
Biogenesis of this superfamily of more than 30 pili requires outer membrane pro-teins known as ushers working together with periplasmic chaperones. P-pili repre-sent the prototype organelle assembled by the chaperone±usher pathway, are ex-pressed by many strains of uropathogenic E. coli, have been shown to be requiredfor the establishment of pyelonephritis and are encoded by the pap (pyelonephritisassociated pili) operon. Each pilus consists of a thick, rigid rod with a thinner,more flexible tip fibrillum at its distal end. The rod consists of PapA subunits ar-ranged to form a hollow, right-handed helical structure 6.8 nm in diameter, whilethe tip fibrillum is composed mainly of repeating subunits of the PapE protein ar-ranged in an open-helical fiber 2 nm in diameter. The PapG adhesin is situated atthe distal end of the tip fibrillum and binds to Gala(1±4)Gal epitopes present inthe globo series of glycolipids found in the human kidney. The PapF and PapKsubunits are present in very low copy number in the pilus and connect the tip fi-brillum to PapG. Assembly of the P-pilus involves two dedicated proteins, the peri-plasmic chaperone PapD and the outer membrane usher protein PapC and is de-scribed under Section 7.2.2.
Type 4 Pili Biogenesis
This pathway for assembling adhesive pili is exemplified by type 4 or bundle-forming pili expressed by a number of Gram-negative bacteria. These long, po-larly localized pili are responsible for twitching motility and bacterial aggregation.Assembly requires 14 or more components, which are thought to organize into asupramolecular structure involved in secretion. Translocation of type 4 pili to thecell surface requires an outer membrane protein termed secretin forming large oli-gomeric rings with apparent central pores. The secretin plays a role analogous tothat of the usher protein.
1 Structure of the Bacterial Cell24

Curli Biogenesis
The third pathway assembles fibers known as curli or thin aggregative pili. Curliare thin filaments expressed by E. coli and Salmonella that can assemble extracellu-lar amyloid fibers 4±6 nm wide. These fibers form a tangled extracellular matrixthat connects several neighboring bacterial cells into small groups (Fig. 1.6).
Curli fibers are thin aggregation surface fimbriae, which are involved in cell±cell attachment and adhesion to extracellular matrices. They are also able to bindhost proteins and might influence the host immune responses. Curli fibers resistprotease digestion and remain insoluble when boiled in 1% sodium dodecyl sul-fate. At least five proteins in E. coli are involved in assembling curli on the cell sur-face (see Section 8.6.2). Regulation of transcription of both operons is complexand is subject to different environmental cues, including osmolarity, temperature,growth phase and protein aggregation in the periplasm. The csgBA operon codesfor the two homologous proteins, CsgA and CsgB, that are secreted into the extra-cellular environment. While the 13-kDa CsgA protein is the major component ofthe curli fiber, CsgB, the minor curli subunit, is required for CsgA polymerizationat the cell surface. It has been suggested that CsgB induces a conformationalchange in CsgA that nucleates its assembly into fibers. CsgA can be purified in asoluble, unassembled state, which, after prolonged incubation, spontaneously as-
1.7 Appendages 25
Fig. 1.6 High resolution deep-etch EM micrograph of curliated E. coli cells.(A±C) The curli fiber meshwork surrounding E. coli cells. (D) Purified CsgAmajor curli can form amyloid fibers by in vitro polymerization. M.R. Chapman,et al. 2003, ASM News 69, 121±126; Fig. 1.

sembles into amyloid fibers. Many amyloid proteins assemble by nucleation-de-pendent polymerization, where the rate-limiting step is nucleus formation.
Why do bacteria produce amyloid fibers? Curli proteins are important media-tors of biofilm formation and are able to bind to eukaryotic extracellular matrixproteins, such as fibronection and laminin. In addition, human macrophages re-cognize and respond better to curliated bacteria than to those not producing thisextracellular matrix. Bacterial amyloids could also play a direct role in certain hu-man neurodegenerative diseases by inducing host proteins to form pathogenicamyloid fibers. Such a cross-species nucleation has been demonstrated amongprions from different genera of yeast.
T-Pilus Biogenesis
A. tumefaciens is able to induce tumors in plants (see Section 10.2.1), and pili (theT-pili) play a key role in virulence. One T-pilus is formed at one end of the cellwhen A. tumefaciens cells perceive phenolic compounds that signal the expressionof virulence genes (see Section 6.2.3) which are located on a plasmid termed theTi-plasmid (tumor-inducing). The products of the virB operon, comprising a totalof 11 genes, are essential for the biogenesis of the T-pilus, which is mainly gener-ated at one end of the bacterial cell. T-pili do not retract but wind into compactcoils which presumably brings the bacterium and host cell into close proximity.In addition to the T-pilus, a number of flagella and common pili of3±4 nm diameter are produced, arranged circumthecally and involved in motility.The common pili are encoded by the chromosome and their function remains elu-sive. The T-pilus serves as a conduit with its 2-nm width lumen for folded pilussubunits, the proteins VirB5 and VirE2 and the single-stranded T-DNA-VirD2complex.
The T-pilin, 74 amino acids in length and product of the virB2 gene, is ligatedbetween its N- and C-termini by a peptide bond, forming a cyclic peptide (see Sec-tion 6.5.5) which is highly resistant to various chemical treatments including gly-cerol; and sodium dodecyl sulfate fails to dissociate T-pili. The remaining tengenes of the virB operon are thought to make up the T-DNA transmembranetransport apparatus, as shown in Fig. 1.7. This transport apparatus appears to alsobe used to deliver T-pilin subunits across the membranes to the outside of the cellwhere they assembly into the pilus. While VirB6, VirB7, VirB8, VirB9 and VirB10are components of the transporter, VirB4 and VirB11 possess ATPase activity,which could be used to promote the transport of the T-DNA complex and the T-pi-lin subunits. VirB1*, the processed form of VirB1, is present in the culture super-natant and is thought to aid transiently in the translocation of T-pilin subunits. Itcould be released when T-pilin subunits reach the outer bacterial cell surface.VirB5 cofractionates as a minor component with T-pili preparations and could sta-bilize T-pilin multimerization and assembly into the T-pilus filament. A potentialprerequisite for T-pilus formation is the hydrolysis of peptidoglycan. The VirB1protein, a transglycosidase, might be involved in peptidoglycan hydrolysis, allow-ing the T-pilus to assemble and penetrate through the peptidoglycan layer.
1 Structure of the Bacterial Cell26

1.7 Appendages 27
Fig. 1.7 T-pilus biogenesis. A transmembrane transport apparatus is compo-sed of VirB proteins that provide a scaffold for the assembly of T-pilin subunits.The pilus itself consists of circular T-pilin subunits with some attached VirB5subunits. E. Lai, C.I. Kado 2000, Trends Microbiol. 8, 361±369; Fig. 5.