1 Digestion in Teleost Fishes_2

of 23
-
Upload
evi-nurul-ihsan -
Category
Documents
-
view
220 -
download
0
Transcript of 1 Digestion in Teleost Fishes_2
-
8/6/2019 1 Digestion in Teleost Fishes_2
1/23
CHAPTER I.
Digestion in Teleost Fishes
L. S. Smith
University of Washington
Seattle, Washington
1. INTRODUCTION
2. OVERVIEW
3. ANATOMY AND GENERAL PHYSIOLOGY OF THE GUT
4. CHARACTERISTICS OF ENZYMES AND OTHER DIGESTIVE SECRETIONS
5. METHODS OF MEASUREMENT AND ANALYSIS FOR DIGESTION STUDIES
6. DIGESTIVE ANATOMY OF SELECTED FISHES
7. REFERENCES
http://www.fao.org/docrep/x5738e/x5738e02.htm#1.%20introductionhttp://www.fao.org/docrep/x5738e/x5738e02.htm#2.%20overviewhttp://www.fao.org/docrep/x5738e/x5738e02.htm#3.%20anatomy%20and%20general%20physiology%20of%20the%20guthttp://www.fao.org/docrep/x5738e/x5738e02.htm#4.%20characteristics%20of%20enzymes%20and%20other%20digestive%20secretionshttp://www.fao.org/docrep/x5738e/x5738e02.htm#5.%20methods%20of%20measurement%20and%20analysis%20for%20digestion%20studieshttp://www.fao.org/docrep/x5738e/x5738e02.htm#6.%20digestive%20anatomy%20of%20selected%20fisheshttp://www.fao.org/docrep/x5738e/x5738e02.htm#7.%20referenceshttp://www.fao.org/docrep/x5738e/x5738e02.htm#1.%20introductionhttp://www.fao.org/docrep/x5738e/x5738e02.htm#2.%20overviewhttp://www.fao.org/docrep/x5738e/x5738e02.htm#3.%20anatomy%20and%20general%20physiology%20of%20the%20guthttp://www.fao.org/docrep/x5738e/x5738e02.htm#4.%20characteristics%20of%20enzymes%20and%20other%20digestive%20secretionshttp://www.fao.org/docrep/x5738e/x5738e02.htm#5.%20methods%20of%20measurement%20and%20analysis%20for%20digestion%20studieshttp://www.fao.org/docrep/x5738e/x5738e02.htm#6.%20digestive%20anatomy%20of%20selected%20fisheshttp://www.fao.org/docrep/x5738e/x5738e02.htm#7.%20references -
8/6/2019 1 Digestion in Teleost Fishes_2
2/23
1. INTRODUCTION
Literature about digestive functions in teleost fishes is extensive in some areas, almost
nil in others. The material to be presented here is not a comprehensive review of that
literature because there are two recent reviews (Harder, 1975; Kapoor et al., 1975),
which have extensive bibliographies on both the anatomy and physiology of teleost
digestive systems. Readers desiring additional information should consult both reviews
because they tend to be complementary, with Harder emphasizing anatomy of European
fishes while Kapoor and co-authors emphasize digestion in North American and Asian
fishes.
The general organization of this chapter begins with the anatomy of the gut, proceeding
from anterior to posterior. Then the route is retraced to discuss the organ physiology of
the digestive tract. A third pass through the tract discusses details of the enzymes. Thenthe typical methods used for studying digestion are described as a basis for the reader to
make detailed comparisons among different methodologies and specific results or to
perform his own experiments. Finally, some anatomical and functional comparisons are
made for four species of fish with distinctly different feeding habits: carp (plant-
oriented omnivore), catfish (animal-oriented omnivore), milkfish (specialized
microplanktivore), and rainbow trout (carnivore). These comparisons are best made
when accompanied by dissections of specimens of each species, although drawings are
provided for readers having no access to the fish.
2. OVERVIEW
2.1 Definition of the Gut and its Subdivisions
2.2 Evolution and Ontogeny of the Digestive Tract
2.3 Generalizations
2.1 Definition of the Gut and its Subdivisions
The gut is a tubular structure beginning at the mouth and ending at the anus. It is
commonly divided into four parts. The most anterior part, the head gut, is most oftenconsidered in terms of its two components, the oral (buccal) and gill (branchial,
pharyngeal) cavities. The foregut begins at the posterior edge of the gills and includes
the oesophagus, the stomach, and the pylorus. In fish, such as the cyprinus, which lack
both a stomach and pylorus, the foregut consists of the oesophagus and an intestine
anterior to the opening of the bile duct. This posterior demarcation is arbitrary and
primarily for convenience during gross dissection and may have little relation to the
functional aspects. The midgut includes the intestine posterior to the pylorus, often with
no distinct demarcation posteriorly between it and the hindgut. The midgut often
includes a variable number of pyloric caecae (pyloric appendages) near the pylorus,
although pyloric caecae are always absent in fishes which lack stomachs. The midgut is
always the longest portion of the gut and ma be coiled into complicated loops (often
http://www.fao.org/docrep/x5738e/x5738e02.htm#2.1%20definition%20of%20the%20gut%20and%20its%20subdivisionshttp://www.fao.org/docrep/x5738e/x5738e02.htm#2.2%20evolution%20and%20ontogeny%20of%20the%20digestive%20tracthttp://www.fao.org/docrep/x5738e/x5738e02.htm#2.3%20generalizationshttp://www.fao.org/docrep/x5738e/x5738e02.htm#2.1%20definition%20of%20the%20gut%20and%20its%20subdivisionshttp://www.fao.org/docrep/x5738e/x5738e02.htm#2.2%20evolution%20and%20ontogeny%20of%20the%20digestive%20tracthttp://www.fao.org/docrep/x5738e/x5738e02.htm#2.3%20generalizations -
8/6/2019 1 Digestion in Teleost Fishes_2
3/23
characteristic for each species) when longer than the visceral cavity. In some fish, the
beginning of the hindgut is marked by an increase in diameter of the gut. The posterior
end of the hindgut is the anus. Only rarely is there a hindgut caecum in fish comparable
to that found in mammals. A cloaca (a chamber common to anal and urogenital
openings and formed from infolded body wall) never occurs in teleost fish, except theDipnoi, although it is universal in sharks and rays.
2.2 Evolution and Ontogeny of the Digestive Tract
The gut of protochordates consists of a simple, straight tube through which food is
propelled by ciliary action. An early elaboration of the gut is seen in lampreys where an
infolding (typhlosole) of the gut wall presumably increases the absorptive area of the
gut. A similar, but spiral, infolding of the hindgut occurs in sharks, rays, and the
coelocanth (Latimeria) in the form of the spiral valve (spiral intestine). The gut wall in
lampreys also contains diagonal muscle fibres, although true peristalsis (travelling wave
of contraction) is thought not to occur. Teleost fish have a gut which is typical of the
higher vertebrates in many respects, although the midgut villi (absorptive papilli) of
mammals are absent in fish.
The gut forms very early during embryological development (ontogeny) and shows
some of the same stages of development as in the evolution of the vertebrate gut, some
larval fish having portions of their gut which are ciliated, for example. The general
character and even the length of the gut may change during development. The gut
appears to shorten, for example, in fish in which the larval stage is herbivorous and the
adult stage is carnivorous. In other fish the gut length remains relatively constant in
proportion to body size throughout life.
2.3 Generalizations
A number of generalizations about the gut of fishes have been attempted, many of them
extrapolated from terrestrial vertebrates. The commonest of these, the observation that
herbivores have longer guts than carnivores, appears only partially true in fish. While
this may betrue in limited groups of fish, it is not universal in teleosts as a whole. Gut
lengths have been listed as 0.2-2.5, 0.6-8.0 and 0.8-15.0 times body length in
carnivores, omnivores, and herbivores, respectively. Thus, the longest guts are found in
herbivores, but not all herbivores have long guts; i.e., the gut lengths of some herbivores
are shorter than those of some carnivores. Part of the explanation lies in the fact thatmany fish eat a variety of food, sometimes ingested with considerable indigestible
material (e.g. mud) which often influences gut length. The size of the food particles -
from submicroscopic plankton to whole fish - may also influence gut configuration.
One generalization so far appears to have no exception. In fishes having no stomachs,
no acid phase of digestion occurs, even when the midgut develops stomach-like pouches
anteriorly. Although gut tissues exhibit great versatility, the midgut appears unable (or
does not need) to duplicate the stomach functions.
In general, most studies relating food habits to gut morphology show considerable
relationship between the two. However, the gut also retains considerable reserve ability
-
8/6/2019 1 Digestion in Teleost Fishes_2
4/23
to respond to new foods, new environments, and new opportunities. This versatility has
been demonstrated in a number of cases in which a single genus has adapted to new
niches and evolved whole new modes of feeding and digestion to utilize otherwise
unexploited food resources and done so over rather short evolutionary periods of time.
At the same time, there are usually severe constraints on adaptations to new food. As
long as swimming continues to be important to a fish's lifestyle, any major change in
body shape, such as a bulging visceral mass resulting from enlarging the stomach or
lengthening the midgut, must extract a penalty in terms of increased effort needed for
swimming. Feeding mechanisms must not interfere with the respiratory functions of the
gills and vice versa. All in all, "packaged" so that any major change in the digestive
system would call for major compromises in many other systems. Perhaps the best
generalization is that teleost fish maintain an intimate relationship between the form and
function of their gut and their food resource. In the final analysis, all of the other life
processes continue to function only when sufficient materials and energy are obtained
and assimilated via the gut.
3. ANATOMY AND GENERAL PHYSIOLOGY OF
THE GUT
3.1 Functional Anatomy of the Gut3.2 Peristalsis and its Control
3.3 Gastric Evacuation Time and Related Studies
3.4 Digestion and Absorption
3.5 Specific Dynamic Action (SDA)
3.6 Interrelationship between Osmoregulation and Digestion
3.1 Functional Anatomy of the Gut
The mouth exhibits a variety of fascinating adaptations for capturing, holding and
sorting food, ratcheting it into the oesophagus and otherwise manipulating it prior toentry into the stomach. Only two which have possible relevance to digestion Will be
discussed.
In milkfish (Chanos), the gill cavity contains epibranchial (suprabranchia) organs
dorsally on each side, consisting either of simple blind sacs or elaborate, spirally-coiled
ducts. The organs occur in several relatively unrelated families of lower teleosts and
apparently relate to the kind of food eaten. Those fish with simple ducts all eat macro-
plankton and those with the larger ducts microplankton. Although their function is
unknown, concentrating the plankton has been suggested as a possibility.
http://www.fao.org/docrep/x5738e/x5738e02.htm#3.1%20functional%20anatomy%20of%20the%20guthttp://www.fao.org/docrep/x5738e/x5738e02.htm#3.2%20peristalsis%20and%20its%20controlhttp://www.fao.org/docrep/x5738e/x5738e02.htm#3.3%20gastric%20evacuation%20time%20and%20related%20studieshttp://www.fao.org/docrep/x5738e/x5738e02.htm#3.4%20digestion%20and%20absorptionhttp://www.fao.org/docrep/x5738e/x5738e02.htm#3.5%20specific%20dynamic%20action%20(sda)http://www.fao.org/docrep/x5738e/x5738e02.htm#3.6%20interrelationship%20between%20osmoregulation%20and%20digestionhttp://www.fao.org/docrep/x5738e/x5738e02.htm#3.1%20functional%20anatomy%20of%20the%20guthttp://www.fao.org/docrep/x5738e/x5738e02.htm#3.2%20peristalsis%20and%20its%20controlhttp://www.fao.org/docrep/x5738e/x5738e02.htm#3.3%20gastric%20evacuation%20time%20and%20related%20studieshttp://www.fao.org/docrep/x5738e/x5738e02.htm#3.4%20digestion%20and%20absorptionhttp://www.fao.org/docrep/x5738e/x5738e02.htm#3.5%20specific%20dynamic%20action%20(sda)http://www.fao.org/docrep/x5738e/x5738e02.htm#3.6%20interrelationship%20between%20osmoregulation%20and%20digestion -
8/6/2019 1 Digestion in Teleost Fishes_2
5/23
The common carp provides an excellent example of non-mandibular teeth being used as
the primary chewing apparatus. Pharyngeal teeth occur in the most fully developed
forms of the Cyprinidae and Cobitidae, although many other groups also show some
degree of abrading or triturating ability with some part of the gill bars. In carp, the
lower ends of the gill bars have a well developed musculature which operates two setsof interdigitating teeth so as to grind plants into small pieces before swallowing them.
The grinding presumably increases the rather small proportion of plant cells which can
otherwise be successfully attached by digestive enzymes.
Many fish which chew their food have some ability to secrete mucus at the same time
and place. This would have some apparent benefit when ingesting abrasive food.
Although one might be tempted to equate such secretions with saliva, enzyme activity
in the mucus does not appear to have been demonstrated, so the mucus is only partly
comparable to saliva.
The oesophagus, in most cases, is a short, broad, muscular passageway between themouth and the stomach. Taste buds are usually present along with additional mucus
cells. Freshwater fishes are reputed to have longer (stronger?) oesophageal muscles than
marine fish, presumably because of the osmoregulatory advantage to be gained by
squeezing out the greatest possible amount of water from their food (i.e., marine fish
would be drinking seawater in addition to that ingested with their food and freshwater
fish would have to excrete any excess water).
The oesophagus of eels (Anguilla) is an exception to this general pattern. It is relatively
long, narrow, and serves during seawater residence to dilute ingested seawater before it
reaches the stomach. A possible conflict between the osmoregulatory and digestive
roles of the gut in marine fish in general will be discussed later (Section 3.5).
Fish stomachs may be classified into four general configurations. These include (a) a
straight stomach with an enlarged lumen, as in Esox, (b) a U-shaped stomach with
enlarged lumen as in Salmo, Coregonus, Clupea, (c) a stomach shaped like a Y on its
side, i.e., the stem of the Y forms a caudally-directed caecum, as in Alosa, Anguilla, the
true cods, and ocean perch, and (d) the absence of a stomach as in cyprinids, gobidids,
cyprinodonts gobies, blennies, scarids and many others, some families of which only
one genus lacks a stomach.
The particular advantage of any configuration seems to rest primarily with the stomachhaving a shape convenient for containing food in the shape in which it is ingested. Fish
which eat mud or other small particles more or less continuously have need for only a
small stomach, if any at all. The Y-shaped stomach, at the other extreme, seems
particularly suited for holding large prey and can readily stretch posteriorly as needed
with little disturbance to the attachments of mesenteries or other organs. Regardless of
configuration, all stomachs probably function similarly by producing hydrochloric acid
and the enzyme, pepsin.
The transport of food from the stomach into the midgut is controlled by a muscular
sphincter, the pylorus. The control of the pylorus has not bean demonstrated in fish, but
the best guess at this time is that it resembles that in higher vertebrates. The pylorus is
-
8/6/2019 1 Digestion in Teleost Fishes_2
6/23
developed to various degrees in different species for unknown reasons, in some species
even being absent. In the latter case, the nearby muscles of the stomach wall take over
this function, which may also include a grinding function by the roughened internal
lining. In fish which lack a stomach, the pylorus is absent and the oesophageal sphincter
serves to prevent regress of food from the intestine, i.e., in fish lacking a stomach andpylorus, the midgut attaches directly to the oesophagus.
The digestive processes of the midgut have not been studied extensively, except histo-
chemically (see Section 4 for details on enzymes), but so far as known resemble the
higher vertebrates. The midgut is mildly alkaline and contains enzymes from the
pancreas and the intestinal wall, as well as bile from the liver. These enzymes attack all
three classes of foods - proteins, lipids, and carbohydrates - although predators such as
salmonids may be largely deficient in carbohydrases. The pyloric caecae attached to the
anterior part of the midgut have attracted considerable attention because of their
elaborate anatomy and their taxonomic significance. Histological examination has
proved them to have the same structure and enzyme content as the upper midgut.Another suggestion was that pyloric caecae might contain bacteria which produce B-
vitamins as in the rodent caecum. When tested, this hypothesis had no factual basis
either. Pyloric caecae apparently represent a way to increase the surface area of the
midgut and nothing more. This still leaves an interesting question of how food is moved
into and out of the blind sacs which are often rather lone and slim: e.g., in salmonids.
The demarcation between midgut and hindgut is often minimal in terms of gross
anatomy, but more readily differentiated histologically - most secretory cells are lacking
in the hindgut except for mucus cells. The blood supply to the hindgut is usually
comparable to that in the posterior midgut, so presumably absorption is continuing
similarly as in the midgut. Formation of faeces and other hindgut functions appear to
have been studied minimally, except histologically.
3.2 Peristalsis and its Control
Peristalsis consists of a travelling wave of contraction of the circular and longitudinal
layers of muscle in the gut wall such that material inside the gut is moved along. The
pharmacology of this system has been investigated in isolated trout intestine
demonstrating that an intrinsic nerve network exists to control peristalsis; i.e.,
cholinergic drugs stimulated and adrenergic drugs inhibited peristaltic movements. The
oesophagus arid stomach are also innervated extrinsically by branches of the vagal(cranial X) nerve. No studies appear to have been made so far concerning details of
food transport through the teleost gut except for measurements of gastric evacuation
time and total food passage time, although gut stasis has been hypothesized to occur in
the Pacific salmon, as in domestic animals.
3.3 Gastric Evacuation Time and Related Studies
Many studies have been performed relating to developing an optimum feeding schedule,
mostly for salmonids, but also including a number of other cultured fish. Variables
considered with feeding rate and gastric evacuation time included temperature, season,
activity, body size, gut capacity, satiety, and metabolic rate. A relatively consistent
-
8/6/2019 1 Digestion in Teleost Fishes_2
7/23
-
8/6/2019 1 Digestion in Teleost Fishes_2
8/23
digestion in channel catfish by Smith and Lovell (1973) showed continuing digestion
(and absorption) of protein during passage through each part of the gut (Table 1). The
methods employed in this study are discussed in Section 4 below. The comparison of
faeces collected from the rectum and from the water also points out the hazard of
incomplete recovery of faecal matter being likely when collection is done from outsidethe gut. Most of the protein digestion occurred in the stomach, but also continued in the
intestine.
Table 1 - Apparent Digestibility of Protein by Channel Catfish 1/
Feed Stomach Upper intestine Lower intestine Rectum Trough
20% protein 61.6 65.4 75.0 80.9 96.7
40% protein 61.4 72.2 86.5 96.6 98.31/ from Smith and Lovell, 1973
Temperature and pH play major roles in determining the effectiveness of digestive
enzymes as a whole (details for specific enzymes are given in Section 4 below).
Although most enzyme production decreases at temperatures above or below
acclimation temperature, most enzyme activity (for a given amount of enzyme)
increases in proportion to the temperature over a wide range of temperatures.
In general, enzyme reaction rates continue to increase at higher temperatures, even
though the temperatures increase beyond the lethal temperatures for the species, until
the enzymes begin to denature around 50-60C. On the other hand, enzymes have
limited ranges of pH over which they function, often as little as 2 pH units. Data forchannel catfish are probably representative of many teleosts. Acid concentrations (pH)
in the stomach ranged from 2 to 4, then became alkaline (pH = 7-9) immediately below
the pylorus, decreased slightly to a maximum of 8.6 in the upper intestine, and finally
neared neutrality in the hindgut (Page et al., 1976). Fish having no stomach have no
acid phase in digestion.
The site of secretion in teleost stomachs appears to be a single kind of cell which
produces both HCl and enzyme(s). This contrasts with mammals where two types of
cells occur, one for acid and one for enzymes. The production of acid in teleosts is
presumably the same as in mammals - NaCl and H2CO3 react to produce NaHCO3 and
HCl, with the blood being the source of both input materials, which are later mostlyreabsorbed in the intestine. One possible explanation for the loss of stomachs in some
species of fish is that they live in a chloride-poor environment and that providing large
amounts of chloride ion for operating a stomach is bioenergetically disadvantageous. In
addition to acid and enzymes, the stomach wall also secretes mucus to protect the
stomach from being digested. As long as the rate of mucus production exceeds the rate
at which it is washed and digested away, the gut wall is protected from being digested.
When mucus production slows or fails, e.g. during gut stasis, during stressful
conditions, or post mortem, the gut wall can be eroded or even perforated by the gut's
own digestive enzymes.
-
8/6/2019 1 Digestion in Teleost Fishes_2
9/23
Two sites produce enzymes in the midgut - the pancreas and the intestinal wall. The
intestinal wall is folded or ridged in simple patterns which can be species specific.
Secretory cells for both mucus and all three classes of enzymes develop in the depths of
the folds, migrate to tops of the ridges (closest to the gut lumen), and then discharge
their products. The pancreatic cells produce enzymes and an alkaline solution which aredelivered to the upper midgut through the common bile duct. The control of pancreatic
secretions (and the pyloric sphincter) in fish is probably the same as in mammals, but
there is no information on teleosts yet.
The physical state of food passing through the gut varies with species and type of food.
Fish, such as salmonids, which eat relatively large prey, reduce the prey in size layer by
layer. Gastric digestion proceeds in a layer of mucus, acid, and enzyme wherever the
stomach wall contacts the food. Food appears liquified only in the midgut and solidifies
somewhat again during formation of faeces. Pellets of commercial feed seem to be
treated similarly, i.e., pellets get smaller and smaller in size with time, although
stomachs of some recently-fed salmonids have been found to contain moderate amountsof liquified pellets. Stomachs of juvenile Pacific salmon captured in the open sea
contained a thick slurry of pieces of amphi-pods in various stages of solubilization. Fish
whose food contains high levels of indigestible ballast, e.g., common carp feeding on a
mixture of mud and plants, probably show minimal change in the appearance or volume
of their food while it passes through the gut. Microphagous fish, such as the milkfish
(Chanos) whose food starts out as a suspension of fine particles, probably also keep it in
much the same form all the way through the gut. In general: there seems not to be the
same degree of liquifaction of food in fish as is commonly described for mammals.
Absorption of soluble food could begin in the stomach - it occurs in mammals, but has
not been investigated in fish - but takes places predominantly in the midgut and
probably to some degree in the hindgut. The sites and mechanisms of absorption are
largely unstudied, except histologically. Several histologists have identified fat droplets
in intestinal epithelial cells following a lipid-rich meal. Increased numbers of leucocytes
in general circulation following a meal by the sea bream and increased number of fat
droplets in them have been described (Smirnova, 1966). It was hypothesized that
leucocytes entered the gut lumen, absorbed lipid droplets, and then returned to the blood
stream. It is clear that the mammalian type of villi with their lymph duct (lacteal) inside
are absent in fish, although there is some folding and ridging of the gut wall to increase
surface area. Lacteals serve as a primary uptake route in mammals for uptake of
droplets of emulsified lipids (chylomicra). Teleost fish have a lymphatic system whichincludes extensions into the gut wall, but its role in lipid uptake is unknown. Absorption
of amino acids, peptides, and simple carbohydrates have been little studied, but
presumably they diffuse through or are transported across the gut epithelium into the
blood stream. What light microscopists identified as a brush border on the surface of the
epithelial cells facing the gut lumen, has now been clarified with electron-microscopy as
microvilli; i.e., subcellular, finger like projections of the cell membrane whose greatly
increased surface area is probably involved in absorption.
3.5 Specific Dynamic Action (SDA)
-
8/6/2019 1 Digestion in Teleost Fishes_2
10/23
Digested food, particularly proteins, is not fully available to a fish even after it has been
absorbed into the blood stream. Amino acids, if used for building new tissue, could be
used as absorbed. If amino acids are to be oxidized for energy, however, deamination
(removal of the amino group) must occur first - a reaction which requires input of
energy. This process, known as specific dynamic action (SDA), can be measuredexternally in fish as an increase in oxygen consumption beginning soon after ingestion
of food followed by an increase in ammonia excretion.
The proportion of amino acids which get deaminated varies with the food and the fish's
circumstances. Fish which are not growing because of low temperature or have their
ration at maintenance level or below, would deaminate most or all of their amino acids.
Fish kept at high rearing temperatures or at high activity levels and therefore having
very high metabolic rates would do likewise. On the other hand, fish having rapid
growth and high protein intake would deaminate a relatively small proportion of their
digested protein, although the absolute quantity of amino acids deaminated could still be
large enough to produce a relatively large SDA. The energy for deamination need notnecessarily come from amino acids, but will be preferentially taken from carbohydrate
or lipid, if available. Thus, salmonid aquaculturists long ago discovered this "protein-
sparing" action of limited amounts of inexpensive carbohydrate in the diet as a way of
reducing the cost of feed and still achieving a desired level of growth. The protein-
sparing action of lipids appears to have been minimally investigated. One can thus
minimize SDA costs, but not avoid them completely.
3.6 Interrelationship between Osmoregulation and Digestion
Researchers studying osmoregulation and researchers studying digestion have rarelyconsidered each other's data. Marine fishes drink significant amounts of seawater, a
relatively-well buffered solution having a pH of about 8.5, while gastric digestion
requires a pH of 4 or lower in most fish. The amount of HCl required just to acidify the
seawater would be substantial, that is, if the entire stomach gets flooded with seawater.
There are several likely alternatives, however. In fish with Y-shaped stomachs, the
seawater could travel directly from the oesophagus to the pylorus, and traverse only a
small fraction of the stomach surface. If, at the same time, digestion functioned
primarily as contact digestion, then it could be largely separated from osmoregulation.
On the other hand, marine salmon stomachs have been found to be filled with a liquid
slurry which would prevent such separation. In such cases, alternation of digestion and
seawater drinking might be possible, although fish whose stomachs seemedcontinuously filled, and therefore would have no time for drinking, have also been
observed.
The pH of seawater should cause little or no problem with intestinal digestion. Too high
a salt content in the intestine might exceed the operational range of some enzymes and
thus reduce the rate of digestion. However, one of the functions of the stomach (and in
eels, the oesophagus) in osmoregulation is to dilute the incoming seawater until it is
approximately equal to the osmolarity of blood, thus protecting the intestine.
The final osmoregulatory product of the gut is a rectal fluid composed of magnesium
and other divalent ions having about the same total concentration as blood. Preliminary
-
8/6/2019 1 Digestion in Teleost Fishes_2
11/23
data from scale loss studies indicated that death occurred from toxic levels of
magnesium in the blood. A possible cause of the high magnesium is that gut peristalsis
stopped, leaving the rectal fluid to accumulate and the magnesium ions to be reabsorbed
instead of being excreted.
Thus, digestion and osmoregulation are so inter-related that problems in one system
could disrupt the functions of the other. Exactly how fish normally avoid such problems
is largely unknown.
4. CHARACTERISTICS OF ENZYMES AND
OTHER DIGESTIVE SECRETIONS
4.1 Digestion in the Mouth and Oesophagus4.2 Digestion in the Stomach
4.3 Digestion in the Midgut and Pyloric Caecae
4.4 The Role of Bile, Gall Bladder and Liver in Digestion
The ability of any organism to digest a given substance rests predominantly on whether
the appropriate enzyme is present or not and then whether the required conditions for
operation of that enzyme exist or not. The following describes the enzymes and their
requisite conditions according to their location.
4.1 Digestion in the Mouth and Oesophagus
The hard surfaces of the mouths of most teleost; fishes would not lead one to expect any
kind of secretion. However, many fish which chew with pharyngeal teeth or similar
structures also produce mucus while chewing. Tests of this mucus in a few species for
enzyme activity have so far yielded negative results. Likewise, oesophageal mucus
cells, when examined histologically, showed no sign of containing any enzymatic
granules, although there are reports of gastric-like secretory cells in the posterior
oesophagus of a few fish.
4.2 Digestion in the Stomach
http://www.fao.org/docrep/x5738e/x5738e02.htm#4.1%20digestion%20in%20the%20mouth%20and%20oesophagushttp://www.fao.org/docrep/x5738e/x5738e02.htm#4.2%20digestion%20in%20the%20stomachhttp://www.fao.org/docrep/x5738e/x5738e02.htm#4.3%20digestion%20in%20the%20midgut%20and%20pyloric%20caecaehttp://www.fao.org/docrep/x5738e/x5738e02.htm#4.4%20the%20role%20of%20bile,%20gall%20bladder%20and%20liver%20in%20digestionhttp://www.fao.org/docrep/x5738e/x5738e02.htm#4.4%20the%20role%20of%20bile,%20gall%20bladder%20and%20liver%20in%20digestionhttp://www.fao.org/docrep/x5738e/x5738e02.htm#4.1%20digestion%20in%20the%20mouth%20and%20oesophagushttp://www.fao.org/docrep/x5738e/x5738e02.htm#4.2%20digestion%20in%20the%20stomachhttp://www.fao.org/docrep/x5738e/x5738e02.htm#4.3%20digestion%20in%20the%20midgut%20and%20pyloric%20caecaehttp://www.fao.org/docrep/x5738e/x5738e02.htm#4.4%20the%20role%20of%20bile,%20gall%20bladder%20and%20liver%20in%20digestion -
8/6/2019 1 Digestion in Teleost Fishes_2
12/23
Pepsin is the predominant gastric enzyme of all vertebrates, including fish. Optimal pH
for maximal proteolytic activity has been reported for several species, as follows:
(a) pH 2 - pike, plaice
(b) pH 3-4 Ictalurus(c) pH 1.3, pH 2.5-3.5 - salmon, probably similar for tuna (Kapoor et al., 1975)
Peptic activity has been shown in a number of cultures and commercial species
including Anguilla japonica, Tilapia mossambica, Pleuronecthys, both Salmo and
Oncorhynchus species, Ictalurus, Micropterus, Lepomis and Perca. The presence of
pepsin is so universal in vertebrates having stomachs that its presence can be presumed
in fish for which no data is available.
The histochemistry of gastric secretion has been little studied in fish, although there is
agreement on the presence of only one type of secretory cell in fish which stains
positively for indicators of pepsinogen (pepsin precursor) cells. There is some questionwhether there may be more than one pepsin present in some fish, but no
chromatographic or other tests have been done to investigate this. Several attempts have
been made to identify acid-secreting cells, but results were either negative or confusing.
Other gastric enzymes have been proposed, but not firmly identified. Chitinolytic
activity with an optimum at pH 4.5 was claimed for the stomach of Salmo irideus, but in
most cases is probably from exogenous sources. If fish are like higher vertebrates, then
the stomach wall also produces the hormone gastrin which stimulates gastric secretion.
A lipase may also be present.
4.3 Digestion in the Midgut and Pyloric Caecae
There are two sources of enzymes for the midgut - the pancreas and the secretory cells
in the gut wall - with the pancreas perhaps secreting the greater variety and quantities of
enzymes in fish. Because of the variety of enzymes present in different species, there
have been some attempts to correlate enzyme activities with diet. However, these
enzyme studies are fragmentary and histochemical tests are too general. Much remains
to be learned about intestinal digestion in fish.
Trypsin appears to be the predominant protease in the midgut. Since the enzyme
appears not to have been isolated, most authors have just tested for proteolytic activityover the pH range of 7 to 11 and reported their results as tryptic activity. The diffuse
nature of the pancreas in most cases has limited many researchers to making relatively
crude extracts from mixed tissues, hampering localization of the enzyme. Tryptic
activity has been found in four stomachless species in Japan: Seriola, two basses and a
puffer. Since these fish lack pepsin, some such kind of protease in the intestine would
be the primary means of protein digestion. Tryptic activity was found in extracts of both
the pancreas of perch and Tilapia and in intestinal extracts of Tilapia, all having a pH
optimum of 8.0-8.2. Proteolytic activity has been identified in the pyloric caecae and
intestine of rainbow trout. In grass carp, tryptic activity was stronger in the intestine
than in the pancreas. In a mixture of pancreatic and pyloric caecae tissue from chinook
salmon, casein was digested maximally at pH 9. Tryptic activity has also been
-
8/6/2019 1 Digestion in Teleost Fishes_2
13/23
demonstrated in extracts of liver of Several species, probably because in fish having a
diffuse pancreas, pancreatic tissue extends into the liver, around the portal veins, and
around the gall bladder. In several of the cases above, when extracts of pancreas were
mixed with extracts of intestine, the tryptic activity increased ten-fold or more,
suggesting the presence in fish of the enzyme enterokinase in the intestinal wall whichactivates in mammals the pancreatic trypsin as it reaches the intestine.
Additional pancreatic enzymes are involved in midgut digestion, many of them yet to be
discovered. For example, Japanese workers are studying the occurrence and
characteristics of a pancreatic collagenase in several Japanese fishes (Yoshinaka et al.,
1973). There have also been several reports of chitinolytic activity in some fish which
eat crustaceans predominantly. This could also have resulted from bacterial activity.
The occurrence of at least one lipase may be assumed in all fishes and has been
demonstrated for a number of species. In carp and killifish extracts of intestine showed
lipolytic activity. In goldfish, lipase activity occurred in extracts of a mixture of liverand pancreas and in the intestinal contents. Esterase (another lipase) activity has been
found in the liver, spleen, bile, intestine, pyloric caecae and stomach of rainbow trout.
Use of radioisotope-labeled lipids in cod suggested that the cod's lipase acted in the
same manner as mammalian pancreatic lipase, although it was not considered more than
a suggestion that fish lipase is of pancreatic origin. Regardless of origin, some kind of
lipase is essential to fish because fatty acids are essential dietary components for fish.
Carbohydrases have perhaps excited the most interest of all the enzymes, particularly
because salmonids do not handle the large carbohydrate molecules very well, and many
workers wanted to determine the reason. Further, because there are several
carbohydrases, the possibility that different enzyme combinations might show
adaptations to different diets also intrigued some investigators. Also, herbivorous fish
might be expected to have more carbohydrase activity and less tryptic activity than
carnivores or omnivores.
Amylase is a widespread starch-digesting enzyme which occurs in human saliva and in
pancreatic secretions into the small intestine. Amylase activity has been found in
goldfish and bluegill sunfish in extracts of mixed liver and pancreas, oesophagus
(contamination from regurgitated food suggested) and intestine, but not in large-mouth
bass. Similar activity has been seen as well in rainbow trout, perch, Tilapia, Pacific
salmon, cod, common carp, eel, and flounder. In fish with a diffuse pancreas there maybe no pancreatic duct and so amylase activity appears in the bile. In mackerel. Scomber
spp., which have a compact pancreas, the bile had no amylase activity.
Other carbohydrases identified included glucosidases (rainbow trout, chum salmon,
common carp), maltase (common carp, red sea bream, Archosargus, marine ayu,
Plecoglossidae), and sucrase, lactase, melibiase, and cellobiase, all of the latter in
common carp. The hypothesis that carnivores might be deficient in one or more
carbohydrases is largely disproved by the widespread presence of amylase in salmonids
and other predators and by the presence of maltase in sea bream and ayu. The
apparently larger diversity of carbohydrases in common carp than in other fish seems
mostly a lack of information about fish other than carp. The question of whether dietary
-
8/6/2019 1 Digestion in Teleost Fishes_2
14/23
differences influence the kind of enzymes present must remain open but the evidence so
far remains largely negative. However, there seems to be some evidence to show that
the amounts of various enzymes may relate to the diet. Data in Table 2 suggest that
herbivores have de-emphasized the production of proteases compared to the carnivores
and the reverse for carbohydrases.
Table 2 - Relative Activity Levels of Amylase and Trypsin in Selected Cyprinids
(Kapoor et al., 1975)
Fish Feeding habit Amylase activity Trypsin activity Amylase Trypsin
Scardinius herbivorous 1.0 0.4 2.5
Blicca omnivorous 1.1 0.9 1.2
Alburus omnivorous 1.0 0.9 1.1
Aspius carnivorous 0.15 1.2 0.125Cyprinus omnivorous 5.8 1.7 3.4
Similarly, in studies of Trachurus, Scomber, Mullus, Mugil, and Pleuronectes, the
predatory species, Trachurus and Scomber had the highest proteolytic and lipolytic
activities, while the planktivore, Mugil, had the lowest proteolytic and the highest
amylolytic activities. Also, stomachless fish (which lack pepsin) are usually herbivores
or omnivores, while carnivorous fish have true stomachs with peptic digestion. On the
other hand, differences in proteolytic activity between Tilapia and Perca were small,
and some other investigations of a variety of species failed to find any species
differences. Apparently, where fish are somewhat specialized in their diets, differences
in their enzyme activities are apparent. Many fish, however, remain non-specialized and
have diversified diets and enzymes.
4.4 The Role of Bile, Gall Bladder and Liver in Digestion
The functions of bile have scarcely been studied in fish, but presumably resemble those
in higher vertebrates. In mammals bile is composed mainly of bilirubin and biliverdin,
which are breakdown products of haemoglobin, and is produced continuously. These
salts act like detergents and serve to emulsify lipids, thus making lipids more accessible
to enzymes because of the increased surface area, allowing some lipids to be absorbed
undigested as micro-droplets. In mammals, about 80 percent of the bile is recycledthrough the liver and gall bladder.
There are a few studies in fish which suggest that bile serves similar functions in fish.
Several histologists have histochemically identified micro-droplets of lipid in midgut
epithelium of fishes. That the gall bladder in fish reabsorbs water as in mammals has
been confirmed. That bile is produced continuously in fish is suggested by the presence
of green mucus in the lumen of the atrophied gut of spawning salmon. There appear to
be no studies in fish of gall bladder contraction or other mechanisms controlling the
release of bile during digestion. An observation of salmon having impacted gall
bladders seemed related to diet because the gall bladders returned to normal when their
dry pellet diet was changed to a moist pellet. Fish having impacted (and presumably
-
8/6/2019 1 Digestion in Teleost Fishes_2
15/23
non-contractile) gall bladders were normal otherwise and were indistinguishable in
appearance and growth rates from fish in the same population with normal gall
bladders.
Anatomists have tried for many years to correlate the shape of the liver and the positionof the gall bladder in the liver with some of its functions. The basic functions of the
liver in processing the foods which have been digested and absorbed are entirely
cellular and molecular in scope. Thus, there is no functional requirement for shape at
any level above the cellular level; i.e., livers basically could be of any shape. On the
other hand, some restrictions are created by its position in the circulatory system
between the gut and the heart, and the necessary interdigitation of the portal and hepatic
veins, hepatic arteries, and bile ducts, all of which must serve essentially every cell of
the liver. In common carp, the liver seems to have no shape of its own and simply fills
every available space between the loops of the intestine. On the other hand, many fish
(e.g., salmonids) have distinctive shape and colour to their livers. Changes in normal
size and shape can indicate dietary or other problems. For example, a large, yellowishliver, often with white blotches suggests fatty degeneration of the liver caused by too
much starch or by using saturated (mammalian) fats in the diet.
5. METHODS OF MEASUREMENT AND
ANALYSIS FOR DIGESTION STUDIES
5.1 Measurement of Stomach Contents
5.2 Measurement of Digestibility and Related Factors
As great diversity of gut anatomy and function occurs in response to the wide variety of
foods found in nature, so also is there a great variety of methods to study digestion. This
multiple diversity often makes comparisons between species impossible and
comparisons within species using different methods difficult. Although the impact of
methods on interpretation of results is always important in scientific experiments, the
problems of methodology in studying fish digestion seem more severe than in most
experimentation. Further, there are significant gaps in information about fish digestion,
particularly in the areas of mechanisms controlling the gut, which have probably comeabout because of the lack of appropriate methods.
5.1 Measurement of Stomach Contents
The most common method of determining gastric evacuation time and the digestive
action of the stomach has been serial slaughter. This involves feeding a population of
fish to a specified level (usually a percentage of body weight), then killing portions of
the population at various times afterwards and analyzing the remaining stomach
contents (analytic methods discussed below). Problems involved in this method include
the variability of food intake by different individuals and the stress imposed on the
http://www.fao.org/docrep/x5738e/x5738e02.htm#5.1%20measurement%20of%20stomach%20contentshttp://www.fao.org/docrep/x5738e/x5738e02.htm#5.2%20measurement%20of%20digestibility%20and%20related%20factorshttp://www.fao.org/docrep/x5738e/x5738e02.htm#5.1%20measurement%20of%20stomach%20contentshttp://www.fao.org/docrep/x5738e/x5738e02.htm#5.2%20measurement%20of%20digestibility%20and%20related%20factors -
8/6/2019 1 Digestion in Teleost Fishes_2
16/23
population by capturing sub-samples: chasing and fright would be expected to inhibit
digestion.
There are a number of variations on this basic technique. In one of these, sockeye
salmon were frozen immediately after sampling, and the stomach contents removed as asingle frozen mass as the fish thawed, thus enabling a complete and reproducible
removal of the contents of a specifically delimited part of the gut. Proximate analysis
was then performed on the stomach contents. Several investigators have devised
stomach pumps for several species of fish. Most commonly it consists of a plastic
syringe of a diameter to fit the oesophagus with the lower (needle) end cut off to give an
opening the same size as the bore. The syringe is inserted under anaesthesia and a
sample of food drawn from the stomach into the syringe. This provides a sample of food
for qualitative, rather than quantitative, analysis for determining enzyme action,
acidification rates, etc. For measuring gastric evacuation in predatory fish, several
investigators have fed whole, smaller fish (pre-weighed), then put the predators into
narrow holding chambers (to reduce the water thickness), and followed the digestion ofthe prey by watching the gradual disappearance of the prey's skeleton by making X-ray
plates. The advantages are: not having to handle the fish to make measurements, and
obtaining repeated measurements from the same fish. Several investigators have added
inert (non-digestible, non-absorbable) substances to commercial diets and measured
periodically the amount remaining in the stomach. Substances used include chromium
oxide (Cr2O3) and radio-isotopes such as cerium-141.
A variety of inert materials have been inserted into fish stomachs to obtain samples of
gastric juices. Spongey foam plastic has been inserted under anaesthesia and removed
later. The contents of the sponge were then squeezed out and analyzed for acid and
enzyme content. Glass beads were inserted into the fish stomachs to test the effects of
stomach distension. A disadvantage of both methods was that the inserted materials
were much easier to acidify (not as well buffered) than food, and that the stomach may
not have responded to them in typical fashion.
5.2 Measurement of Digestibility and Related Factors
Chromium oxide (Cr2O3) mixed with prepared diets and measured in the faeces provides
a general comparison of the overall digestibility of a feed expressed as:
More specific measurements of digestibility now seem to be replacing the use of Cr2O3as an indicator. Measurements of the caloric values of ingested food and faeces
produced provides some part of the information for estimating the energy balance for a
fish (oxygen consumption and growth rate are also needed). Alternatively, protein
(measured as nitrogen content) or lipid content of food and faeces could be measured. In
both cases the equation would be:
-
8/6/2019 1 Digestion in Teleost Fishes_2
17/23
In measuring protein nitrogen, one really should take the nitrogen from the deaminated
amino acids into account, which requires measuring the ammonia excreted by the gills(metabolic nitrogen). That equation should be:
Some of the most extensive studies on the digestibility of food components have been
performed and reported by Phillips (1969) and readers should see these for detailed
methods.
Several authors have combined the use of an indicator or other general measurement
with measurement of one food component, generally protein. As an example. Smith and
Lovell (1973) combined measurement of protein nitrogen with Cr2O3. Their equation
was:
An earlier author (cited by Kapoor et al., 1975) used the same idea and substituted
caloric values in feed and faeces for the Cr2O3 in the equation above. By comparing the
digestibility of pure protein and of protein in practical diets. Smith and Lovell (1973)
concluded that their combined indicator method gave results for catfish similar to thedigestive coefficients in livestock feeding tables for concentrated feedstuffs.
Many of the methods described so far require the collection of faeces. A great variety of
devices have been designed to do this, most of them producing a place with a low
velocity water flow so that finely particulate faecal matter is not swept away. That this
can be a problem is illustrated by Table 1 of this paper which shows significant
differences between faecal composition in the trough and in the rectum. Post (1965)
designed a holding chamber to reduce this problem by having static water around the
posterior half of the fish and collecting the faeces from this water. Even collecting
faeces from the rectum of fish does not solve all problems of faecal analysis because it
is difficult to determine by their location in the hindgut when water reabsorption is
complete. Thus faecal collection always includes some degree of compromise which
affects the subsequent results and the comparability with other experiments.
The use of Mette's rods (tubes) has been made primarily by Russian investigators. The
rods consist of short sections of glass tubing filled with solidified substrates; such as
gelatin, coagulated egg white, starch paste, etc. The tubes are placed in the gut and
recovered at some later time when the length of the tube emptied of substrate is
measured as both a qualitative and quantitative indicator of digestion. Placing and
recovering such tubes in the stomach is usually no problem, but getting them into the
midgut is more difficult. In studying intestinal functions in carp and European bream
-
8/6/2019 1 Digestion in Teleost Fishes_2
18/23
(another cyprinid), one solution to placing the tubes in position was to cut one of the
long loops of intestine and bring the ends out through the body wall, i.e., surgically
make an intestinal fistula. The gut openings normally kept plugged, were opened to
insert and remove the Mette's rods.
-
8/6/2019 1 Digestion in Teleost Fishes_2
19/23
6. DIGESTIVE ANATOMY OF SELECTED FISHES
6.1 Rainbow Trout (Salmo gairdneri)6.2 Channel Catfish (Ictalurus punctatus)
6.3 Common Carp (Cyprinus carpio)
6.4 Milkfish (Chanos chanos)
The gross anatomy of four cultured fishes, having diverse feeding habits are described
in this section, emphasizing noteworthy structures which show relationships to feeding
and digestion. The line drawings are intended to serve as a general guide for the
examination and dissection of actual specimens and are not intended to show definitive
details of visceral organs. The four species illustrated were selected primarily for variety
of feeding habits and life styles among a dozen or more cultured species with no intent
to indicate their importance or representativeness compared to other culture species.
6.1 Rainbow Trout (Salmo gairdneri)
Rainbow trout are largely carnivorous, but show few anatomical specializations for
capturing and digesting prey. Teeth are simple and small with no other elaboration of
structures to capture, hold, or swallow prey. Salmonids swallow their food whole via a
wide oesophagus into a Y-shaped stomach. Many pyloric caecae branch near the pyloric
end of the midgut, their numbers often being of taxonomic importance among the
various salmonid species. The pancreas is diffusely scattered in the fat and connective
tissue around the pyloric caecae and is not readily visible. The gall bladder extends from
the middle lobe of the liver and the bile duct can usually be traced from there to the
upper midgut in larger specimens. The midgut merges into the hindgut without any
particular demarcation.
Other visceral organs include a thin-walled, nearly transparent swim bladder, the kidney
just dorsal to that and running the full length of the visceral cavity. The kidney covers
the dorsal aorta on the ventral surface of the vertebral column and encloses the posterior
vena cava. The urinary ducts can usually be seen on the ventral surface of the kidney.
They meet somewhat anterior to the posterior end of the kidney and descend as a single
duct around one side of the swim bladder. An expansion of this descending portion of
the urinary duct serves as a urinary bladder. The bladder is connected to the urogenitalpapilla as are testes in mature males. The gonads develop dorso-laterally in the anterior
visceral cavity in both sexes, but the ovaries have no ducts connecting the urogenital
papilla, eggs are simply shed into the visceral cavity. The only major organ remaining
unmentioned is the spleen. In salmonids this lays ventrally, just above the pelvic fins,
attached to the posterior side of the major visceral mass.
In general.) the rainbow trout is representative of most salmonids. It is a relatively
primitive (unspecialized) fish, a typical carnivore with good swimming ability for
capturing prey, a stomach which can easily extend posteriorly for ingesting relatively
large prey, and a short intestine for handling food containing minimal amounts of
http://www.fao.org/docrep/x5738e/x5738e02.htm#6.1%20rainbow%20trout%20(salmo%20gairdneri)http://www.fao.org/docrep/x5738e/x5738e02.htm#6.2%20channel%20catfish%20(ictalurus%20punctatus)http://www.fao.org/docrep/x5738e/x5738e02.htm#6.3%20common%20carp%20(cyprinus%20carpio)http://www.fao.org/docrep/x5738e/x5738e02.htm#6.4%20milkfish%20(chanos%20chanos)http://www.fao.org/docrep/x5738e/x5738e02.htm#6.1%20rainbow%20trout%20(salmo%20gairdneri)http://www.fao.org/docrep/x5738e/x5738e02.htm#6.2%20channel%20catfish%20(ictalurus%20punctatus)http://www.fao.org/docrep/x5738e/x5738e02.htm#6.3%20common%20carp%20(cyprinus%20carpio)http://www.fao.org/docrep/x5738e/x5738e02.htm#6.4%20milkfish%20(chanos%20chanos) -
8/6/2019 1 Digestion in Teleost Fishes_2
20/23
-
8/6/2019 1 Digestion in Teleost Fishes_2
21/23
expansion of a large meal anyway. The remainder of the visceral organs are relatively
unremarkable (Figure 1c).
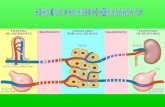
Fig. 1. Diagrammatic representation of the digestive systems of four fish described
in the text, arranged in order of increasing gut length.
a. Rainbow trout (carnivore);
b. Catfish (omnivore emphasizing animal sources food);
c. Carp (omnivore, emphasizing plant sources of food);
d. Milkfish (microphagous planktovore).
6.4 Milkfish (Chanos chanos)
Milkfish are specialized in several respects. The body shape, the streamlined cover over
the eyes, and the widely forked tail all go with a fast-swimming life style. Fine (almost
membranous) gill rakers suggest filter feeding, making a designation as carnivore,
omnivore, or herbivore impossible since plankton is a mixture of many life forms,
including some which are too simple to be clearly plant or animal. Milkfish are most
frequently designated as microphagus planktovores. A specialized epibranchial organ
above and behind the gills may help to concentrate microplankton, although no one has
really demonstrated how it might do so. The stomach is a simple tube, somewhat
convoluted, and of moderate size. The pyloric end of the stomach has thick, muscular
walls and is usually described as a gizzard. The pyloric region of the long, narrow
intestine has numerous pyloric caecae, also of small diameter. The swim bladder and the
lining of the visceral cavity are membranous, similar to those in salmonids except for
being jet black (Figure 1d).
In general, milkfish are cultured in enriched, saltwater lagoons in which they swim
rapidly, straining their food from the typically turbid water.
7. REFERENCES
Harder, W. 1975, Anatomy of fishes. Part I. Text. Part 2. Figures and plates. Stuttgart.
E. Schweizerbart'sche Verlagsbuchhandlung, Pt.1:612 p., Pt.2:132 p. 13 pl.
Kapoor, B.B. 1975, H. Smit and I.A. Verighina, The alimentary canal and digestion in
teleosts. Adv.Mar.Biol. 13:109-239
Magnuson, J.J. 1969, Digestion and food consumption by Skipjack tuna.
Trans.Am.Fish.Soc., 98(3): 379-92
Page, J.W. 1976 et al., Hydrogen ion concentration in the gastrointestinal tract of
channel Catfish. J.Fish Biol., 8:225-8
Phillips, A.M. Jr., 1969 Nutrition, digestion and energy utilization. In Fish physiology,
edited by W.S. Hoar and D.G. Randall. New York, Academic Press, vol. 1:391-432
http://www.fao.org/docrep/x5738e/x5738e13.gifhttp://www.fao.org/docrep/x5738e/x5738e13.gifhttp://www.fao.org/docrep/x5738e/x5738e13.gifhttp://www.fao.org/docrep/x5738e/x5738e13.gif -
8/6/2019 1 Digestion in Teleost Fishes_2
22/23
Post, G., W.E. Shanks and R.R. Smith, 1965 A method for collecting metabolic
excretions from fish. Prog.Fish-Cult. 27:108-88
Smirnova, L.I., 1966 Digestive leukocytosis of bream (Abramis brama). In Biology of
fishes of the Volga reservoirs. Tr.Inst.Biol.Vnutr.Vod./Trans.Inst.Biol.Inland Waters,10(13) -.143-7
Smith, B.W. and R.T. Lovell, 1973 Determination of apparent protein digestibility in
feeds for channel catfish. Trans.Am.Fish.Soc., 102(4):831-5
Yoshinaka, R., M. Sato and S. Ideka, 1973 Studies on collagenase of fish. l. Existence
of colla-genolytic enzyme in pyloric caecae of Seriola quinqueradiata. Bull. Japan. Soc.
Sci.Fish., 39(3):275-81
-
8/6/2019 1 Digestion in Teleost Fishes_2
23/23


![01.3 Digesters Digestion[1]](https://static.fdocuments.net/doc/165x107/577d276f1a28ab4e1ea3eefc/013-digesters-digestion1.jpg)