
1 Animal biotechnology lecture 2 Dr. Ziad Jaradat.
113
1 Animal Animal biotechnology biotechnology lecture 2 lecture 2 Dr. Ziad Jaradat Dr. Ziad Jaradat
-
Upload
melissa-oneal -
Category
Documents
-
view
220 -
download
0
Transcript of 1 Animal biotechnology lecture 2 Dr. Ziad Jaradat.

1
Animal biotechnology Animal biotechnology lecture 2lecture 2
Animal biotechnology Animal biotechnology lecture 2lecture 2
Dr. Ziad JaradatDr. Ziad Jaradat

2
Animal Biotechnology & Transgenic Animals
• Since the early 1980s, fruit flies, fish, sea urchins, frogs, laboratory mice and farm animals, such as cows, pigs, and sheep have been successfully produced.
• The ability to manipulate the genome of the whole animal and the production of transgenic animals has influenced the science dramatically in the last 15 years.
• The procedure for introducing exogenous donor DNA into a recipient cell is called Transfection.
• Chromosomes are taken up inefficiently so that intact chromosomes rarely survived the procedure. Instead the recipient cell usually get a part of the DNA.

3
• Now, with the advent of the recombinant DNA, the possibility of introducing a particular segment of DNA become possible. However, still there are always some problems of the stability of the new inserts (transient transfectants).
• An exciting development of transfection techniques is the application of DNA technology to introduce genes into animals.

4
• An animal that gains new genetic information from the addition of foreign DNA is described as Transgenic while the introduced DNA is called the transgene.
• The transgenes are introduced into the pronuclei of fertilized eggs by injection, and the injected embryos are incubated in vitro or implanted into the uterus of a pseudopregnant female for subsequent development.
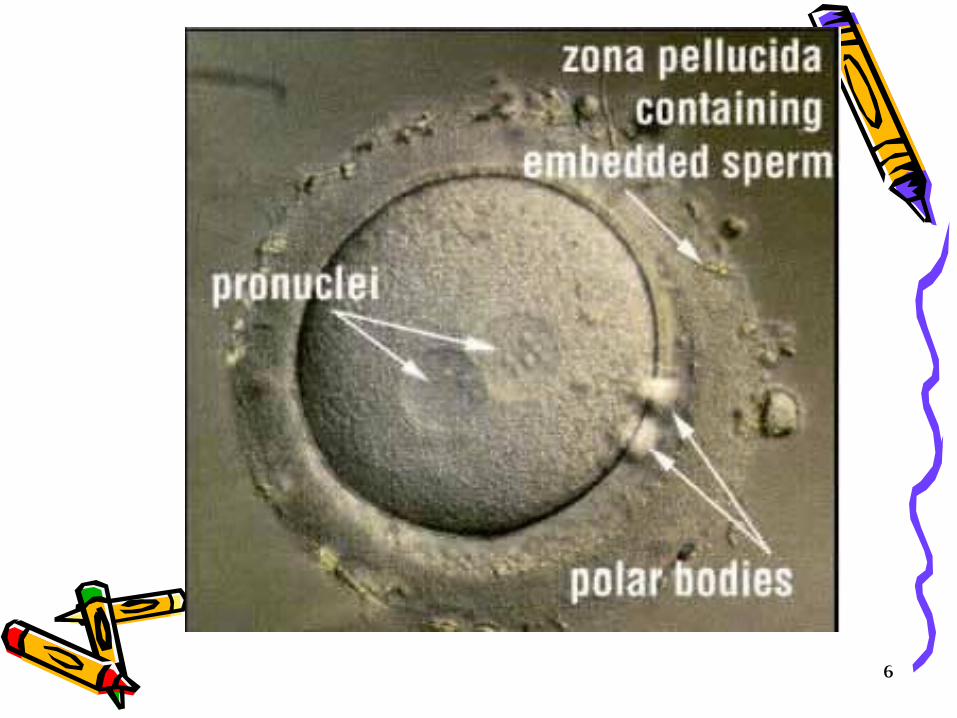
What is Pronucleus? For a short time after fertilization, the male pronucleus and female pronucleus exist separately.
• Female pronucleus; In the maturing of the ovum preparatory to impregnation, a part of the germinal vesicle becomes converted into a number of small vesicles, which aggregate themselves into a single clear nucleus which travels towards the center of the egg and is called the female pronucleus.

5
• Male pronucleus; In impregnation, the spermatozon which enters the egg soon loses its tail, while the head forms a nucleus, called the male pronucleus, which gradually travels towards the female pronucleus and eventually fuses with it, forming the first segmentation nucleus. The male pronucleus is larger than the female’s and can be seen fairly easily under a light microscope.
6

7
Synopsis of the transgenesis process;• Plasmids carrying the gene of interest are injected into
the germinal vesicle (nucleus) of the oocyte or into the pronucleus (before uniting with the gamete) of the fertilized egg.
• The egg is implanted into a pseudopregnant mouse• After birth, the recipient mouse can be examined to see
whether it has gained the foreign DNA and if so whether it is expressed.
• As a result; multiple copies of transgenes are integrated at random locations in the genome of the transgenic individuals.
• The transgenes in many transgenic individuals are also transmitted through the germline to subsequent generations.

8
Note; If the transgenes are linked with functional promoters, expression of transgenes as well as display of change in phenotype is expected in some of the transgenic individuals
• Questions to be asked about any transgenic animal are;
• how many copies it has of the foreign DNA (varies 1-50)

9
• where these copies are located [usually multiple copies are integrated into a tandem array (arranged adjacent to each other) into a single chromosomal site]
• whether they are present in the germ line and inherited in Mendelian manner.
• can the gene be expressed independently? i.e does the regulatory elements function independently
• are transfected genes expressed with the proper developmental specificity?
• A good result if we obtain 15% of the animals to be transgenic.
• In the progeny of the infected animal, the expression of the donor gene is extremely variable and that could be dependent on the place of integration of the new DNA.

10
Transgenesis; Methodology
• Transgenic technology has been developed and perfected in the laboratory mouse. Since the early1980’s hundreds of different genes have been introduced into various mouse strains. These studies have contributed to;
• understanding of gene regulation• tumor development, example introducing oncogenes
and observe the effect

11
• immunological specificity, example producing knockout genes that are responsible for some immunological aspects
• molecular genetics of development
• other biological interests such as examining the possibility of using transgenic animals in the industrial production of human therapeutic drugs.. etc.

12
Methods of gene transfer in animals
For transgenesis, DNA can be introduced into mice by one of the following methods;
• Retroviral vectors that infects the cells of an early stage embryo prior to implantation into a receptive female.
• Microinjection into the enlarged sperm nucleus (the male pronucleus) of a fertilized egg
• Introduction of genetically engineered embryonic stem cells into an early stage developing embryo prior to implantation into a receptive female.
• Transfer of diploid somatic nuclei into an enucleated oocyte.

13
Retrovirus-Mediated Gene Transfer• The most useful vectors for the purpose of gene
isolation are those that lend themselves to the production of libraries consisting of overlapping fragments of genomic DNA, ideally encompassing the entire genome several times.
• Exmaple; bacteriophage λ genomic library of 106 viruses each containing on average 20 Kb of DNA, represents 6-7 copies of the entire mouse genome and the probability that each gene is represented is very high.
• Retroviruses can be used for the transfer of foreign genes into animal genomes.
•

14
• This can best be done at 4-16 cell stage embryos. However, it can be done up to midgestation, but with incomplete infections i.e low infectivity rate.
• Immediately following infection, the retrovirus produces a DNA copy of its RNA genome using its reverse transcriptase.
• Completion of this process requires that the host cell undergoes the S phase of the cell cycle. Therefore, retroviruses effectively transduce only mitotically active cells.
• Modifications to the retrovirus frequently consist of removal of structural genes, such as gag, pol, and env, which support viral particle formation.

15
• Additionally, most retroviruses and complementary lines are ecotropic in that they infect only rodents, such as rats and mice, and rodent cell lines rather than humans.
• The DNA copy of the viral genome, or provirus, integrates randomly into the host cell genome, usually without deletions or rearrangements.
• Because integration is not by way of homologous recombination, this method is not used effectively for site-directed mutagenesis.
• Very high rates of gene transfer are achieved with the use of retroviruses.

16
Vector Origin Insert size rangeMulticopy plasmids multicopy plasmids up to 20 kb
• Lambda vectors Bacteriophage λ up to 30 kb• Cosmid Bacteriophage λ up to 40 kb• P1 artificial chrom Bacteriophage P1 80-90 kb• Bacterial artificial chrom. Large Bacteria plasmid 100-300 kb
(F factor)• Yeast chrom. (YAC) Yeast chromosome 100-1000 + kb
+ means indefinite.
Table of common vectors used for such purpose

17
Disadvantages of this method include:• Low copy number integration.• Additional steps required to produce
retroviruses.• Limitations on the size of the foreign DNA insert
(usually 9 to 15 kb) transferred.• Potential for undesired genetic recombination
that may alter the retrovirus.• High frequency of mosaicism.• Possible interference by integrated retroviral
sequences in transgene expression.

18
• The genome of the retroviral strain can be integrated into the same nucleus as the transgene. This means that the virus itself could be produced by the transgenic organism and create a problem especially if the animal will be used for production of food.
• Also the provirus attracts methylation which possibly in conjugation with other mechanisms disables its expression when it passes through the germ line.
• Due to this, and to the availability of other alternative methods, the retroviral vector method is rarely used for producing transgenic animals that have a commercial potential.

19
DNA Microinjection Method
Because of the disadvantages of the retroviral vectors, microinjection of DNA is currently the preferred method for producing transgenic mice.
• First - you need the gene of interest in the proper form. A linear transgene construct is made, which contains: – the structural gene of interest, with introns – a strong mouse gene promoter and enhancer to allow the gene to be
expressed – vector DNA to enable the transgene to be inserted into host DNA
• The immature female mice will be induced to superovulate by sequential administration of FSH/LH and HCG and mated to fertile males. One-celled embryos are flushed from the oviducts and placed in a drop of medium and viewed by phase-contrast or interference microscopy.

20
This procedure has the following steps; • The number of available fertilized eggs that
are to be inoculated are increased by stimulating donor females to superovulate.
• This can be done by – Giving the mice an initial injection of pregnant
mare’s ( an adult female of horse or related mammal) serum
– Another injection about 48 hours later of human chorionic gonadotropin (hCG). By this protocol the female produces about 35 eggs instead of the normal number of 5-10.

21
• These females are mated, then sacrificed and the fertilized eggs (oocytes) are flushed from their oviducts and recovered.
• Eggs are treated with hyaluronidase to remove adherent follicle cells.
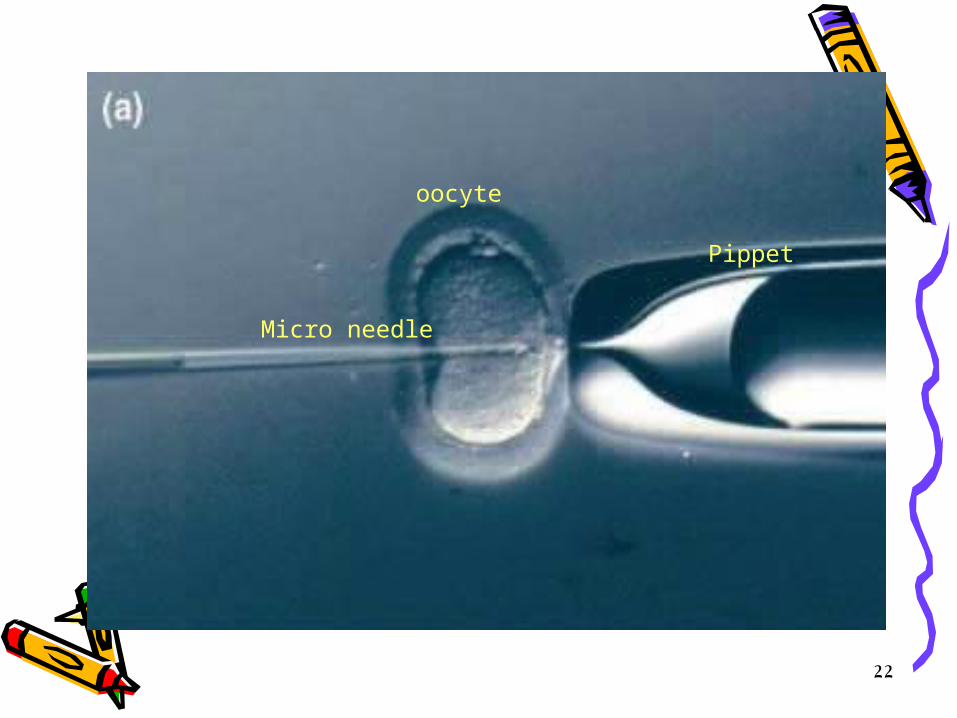
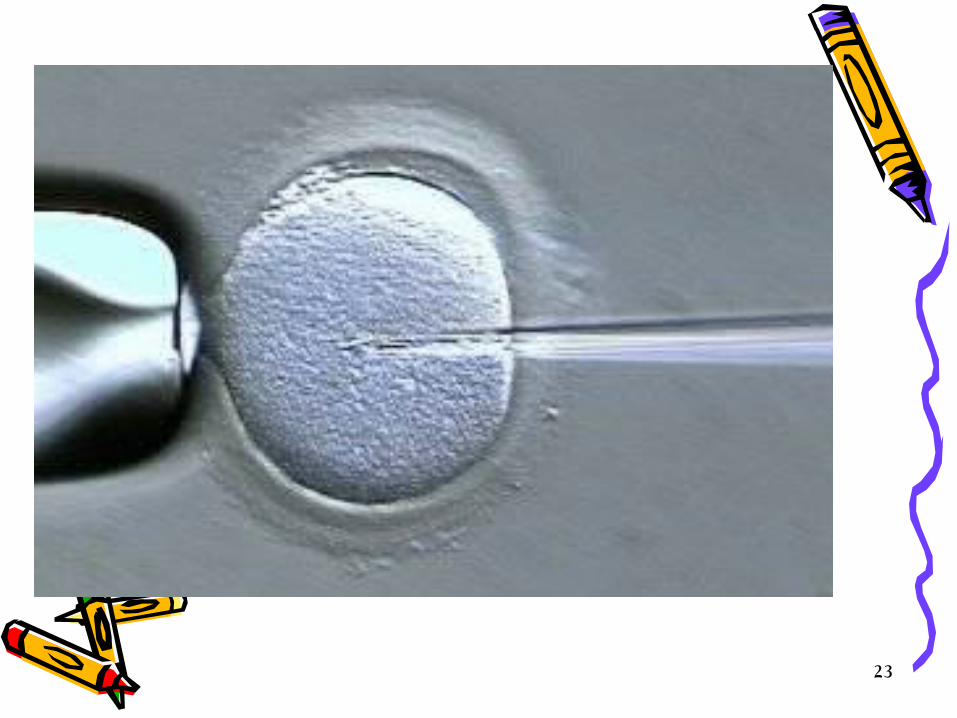
• Unfertilized eggs are discarded • The eggs are inoculated immediately with the transgene, briefly;
– embryo at the pronuclear stage is held in place by suction. – a micro needle loaded with a suspension of plasmid DNA will be
prepared. – It is introduced through the zona pellucida and plasma membrane into
the most accessible pronucleus (usually the male) and – several hundred molecules of the recombinant DNA are injected in a
volume of approximately 1 picoliter (p1). – on a good day several hundred eggs can be injected.
• The male pronucleus can be located by using dissecting microscope and the eggs then can be maneuvered, oriented and held in place while the DNA is microinjected.
22
oocyte
Micro needle
Pippet
23

24
• After inoculation, 25-40 eggs are implanted microscopically into a foster mother who has been made pseudo-pregnant by being mated to a vasectomized male so that none of the eggs of the foster mother will be fertile therefore, the foster mother will deliver pups from the implanted fertile eggs three weeks after the inoculation.
• After birth, the presence of foreign material is studied by DNA hybridization with appropriate probes or PCR.
• A transgenic mouse can be mated to another to produce transgenic homozygous transgenic animal.

25
More details on gene transfer.

Gene Transfer to Animal cells
Dr. Ziad Jaradat

• There are four major strategies for gene transfer to animal cells, two of which are considered biological mechanisms using virus (transduction) or bacterial that invade animal cells (bactofection), those methods involve infection while the other two are chemical and physical methods which do not involve infection thus termed transfection.
• Virus (transduction); the transferred gene represents part of the viral genome
• Bacteria (bactofection); the gene will be transferred as a plasmid

Chemical transfection
• ; DNA will be taken from the surroundings when the DNA is presented as a synthetic complex either as;
– a complex with overall positive charge, allowing it to interact with negatively charge cell membrane and promote uptake by endocytosis
– as lipophilic complex that fuses with the cell membrane and deposits the transgene directly into the cytoplasm.

Physical transfection;
• in this method, naked DNA is deposited directly into the cell by exploiting a physical force. This includes;
• microinjection• particle bombardment• ultrasound • electroporation
• Which ever method used, the result is called transformation which is a change in the recipient cell’s genome caused by the acquired transgene.

Chemical Transfection Techniques
• Calcium phosphate method; involves the formation of a fine DNA/calcium phosphate co-precipitate which first settles on the cells and then internalized by endocytosis.
• The precipitate must be formed freshly at the time of transfection. The DNA escapes and reaches the nucleus and can be then expressed. Since the cells must be coated by the calcium complex, monolayers of cells must be used for maximum efficiency. However, this method gives only 1-2% transfection efficiency.

• Transfection with polyplexes; this is more efficient than that with calcium as it gives more uniform particle size. Polyplexes are a polycationic compounds that form soluble complexes (polyplexes) through spontaneous electrostatic interaction with DNA.
• This method is adapted as a plasmid DNA
transfer. The efficiency of the method can be increased by exposing cells to osmotic shock or treatment of cells with chloroquine.
• Anew generation of the polycationic compounds has been developed such as poly-L-lysine and synthetic poly amines, polyethelylenimines and dendrimers (highly complex molecules built in layers from a central initiator such as ammonia or ethylenediamine).

Transfection with liposomes and lipoplexes
• This can be done by packaging the DNA inside a fusogenic phospholipid vesicle which interacts with the target cell membrane and facilitate DNA uptake.
• Briefly, bacterial cells will be transformed with suitable plasmid vector and treated with chloramphenicol to amplify plasmid copy number.

• The cells will be treated with lysozyme to remove cell wall, resulting in protoplasts that will be centrifuged gently onto a monolayer of mammalian cells to promote fusion among them using polyethylene glycol.
• Using lyposomes is more commonly used for this type of transfection. This method is commonly known as lipofection. This method is far more efficient than chemical transfection method. With this method up to 90% of cells in culture dish can be transected.

Physical transfection methods
• Electroporation Electroporation • Ultrasound Ultrasound
both methods create transient pores in the cellboth methods create transient pores in the cell

Electroporation• is a physical transfection technique involves
creating transient nano-meter size pores in the cell membrane by exposing cells to a brief pulse of electricity. The most critical parameter is the intensity and duration of electrical pulse.
• Electroporation can be used for in vivo gene transfer particularly for surface or near surface tissue such as skin, muscle and certain tumors or even internal tissues such as liver. This can be achieved by direct application of electrodes to the skin following shaving and mild abrasion with the DNA being injected into the skin before electroporation.

Ultrasound transfection
• Involves the exposure of cells to a rapidly oscillating probe such as the tip of sonicator. In this method the application of ultrasound waves to a dish or cells or a particular tissue results in the formation and collapse of bubbles in the liquid, including the cell membrane, a process known as cavitation.
• The transient appearance of such cavities allows the DNA to cross the membrane into the cytoplasm. This method can be used both for in vivo or in vitro as the plasmid DNA is left structurally intact. As for electroporation, the DNA will be injected and then the ultrasound will be applied.

DNA microinjectionDirect transfer of DNA into the cell
without a carrier is called DNA microinjectin. This can only be done for only few cells at a time. This technique is used mainly for large cells such as oocytes, eggs and the cells of early embryos. The DNA can be directly injected into tissues, such as skin, muscle or internal organs or it can be injected into the blood.

38
• The process is remarkably efficient. Up to 60- 66%% of the embryos survive injection and up to 25-30% of the embryos transferred to the oviduct survive to birth and about 25% of pups are transgenic (transgenic founders). Thus, from 1000 inoculated fertile eggs, 30-50 (3-5%) transgenic pups are produced.
• The injected DNA gets incorporated at random sites within the genome and often multiple copies are incorporated at one site, therefore, not all the transgenic animals will have the desired traits.

Particle bombardment• is another direct delivery method
initially developed for the transformation of plants. This method involves coating small metal particles with DNA and accelerating them into target tissues using a powerful force such as the blast of high pressure gas or an electric discharge through a water droplet. In animals, this method is used for tissues such as skin cells in vivo rather than cultured cells.

Bacterial vectors for gene transfer
The exploitation of living bacteria for gene transfer is central to the genetic manipulation of plants. Agrobacterium tumefaciens and its close relatives have been used for 20 years to generate transgenic plants.
• Recently, A. tumefaciens has been used to transfer DNA to human cells. The protoplast fusion technique can be considered as highly efficient form of bactofection. However, this type needs human intervention, while the A. tumefaciens does not require human intervention.

How does the bacterial transfer of DNA happen?
• The bacteria invades the host animal cells and undergo lysis within them releasing their plasmid DNA. Example of these bacteria including Salmonella species (lysis occurs in the phagocytic vesicle), Listeria monocytogenes and Shigella flexneri (lysis occurs for these two species after they escape from the vesicle).

• The plasmid DNA then finds its way to the nucleus where it gets incorporated with the cell’s genome and gets expressed.
• Contrary to the above mentioned bacteria,
A. tumefaciens does not invade the cell, instead, it attaches itself to the outside surface followed by conjugation.

Viruses That are used as gene transfer vectors
• Virus particles have a natural ability to adsorb to the surface of the cells and gain entry. This can be exploited to deliver recombinant DNA into animal cells.
• Several classes of viruses has been used for gene therapy and at least 8 has been used in clinical trials. Transgenes may be incorporated into viral vectors either by addition to the whole genome or by replacing one or more viral genes. This can be done by ligation or by homologous recombination.

• If the transgene replaces a none essential gene the vector is described as helper-independent
• If it replaces an indispensable gene, then this vector will be helper dependent.
• It is generally recommended to use vectors from which all viral coding sequences has been deleted such vectors are described as fully deleted or gutted or gutless vectors. These vectors contain just the cis-acting elements required for packaging and for genome replication.

Advantages of such vectors• high capacity for foreign DNA• because no viral gene products are made, the
vector has no intrinsic cytotoxic effects.
Adeno virus • These are viruses with a linear, double stranded
genome, of approximately 36 kb. These are used frequently due to certain advantages;– stability– high capacity for foreign DNA– wide host range including none-dividing cells– Ability to produce high titer stocks up to 1011 pfu/ml.

Adeno-associate virus (AAV)• These viruses are not genetically related to
adenovirus but is so-called because it was first discovered as a contaminant in an adenovirus isolate.
• The AAV is a single stranded DNA and is a member of the parvovirus family.
• It is naturally replicating deficient, thus it requires the presence of another virus such as adenovirus to complete its infection cycle. AAV replicates lytically and produces thousands of progeny virions.

• The dependence of AAV on a heterologous helper virus such as adeno virus provides an unusual degree of control over vector replication making AAV one of the safest vectors to use for gene therapy.
• Other advantages of this viral vector is the wide host range that it exhibits including none dividing cells.

• The AAV genome is small (5 kb) and comprises a central region containing rep
(replicase) and cap (capsid) genes flanked by 145 kb inverted terminal repeats.
• Foreign DNA replaces the cap region and gets expressed by indigenous promoter. However rep proteins might interfere with the expression process thus responsible for some of the cytotoxic effects of the virus.

• New vectors are designed with the deletion of the rep and cap genes and only utilizing the repetitive sequence which are the only elements required for replication, transcription and proviral integration.
• AAV vectors have been used to introduce genes efficiently into many cells including liver, muscle, and neurons.

Baculovirus vectors (BV)
• Baculovirus promote high level of transgene expression in insect cells but can also infect mammalian cells.
• Have rode shape capsid and large double-stranded DNA genomes. They productively infect arthropods particularly insects.
• BV vectors are used mainly for high–level transient protein expression in insects and insect cells.

• These viruses produce a protein called polyhedron at a very high expression level yet this protein is not necessary for the infection process therefore it can be replaced with foreign DNA that can be expressed at high levels (up to 1 mg / 106 cells) under the control of the endogenous polyhedron promoter.
• The highest amount of expressed proteins
was initially achieved when the transgene was expressed as a fusion protein by incorporating it with the first 30 amino acids of the polyhedron protein.

There are two types of BV that are used as vectors;– Autographa californica multiple
nuclear polyhedrosis virus (AcMNPV); infects mainly insects
– Bombyx mori nuclear polyhedrosis virus (BmNPV); infects insects silkworms and is used for production of recombinant proteins in silk worm larvae.

• One problem of expression of mammalian proteins in insects is the different types of glycosylation. Thus as a solution, is to exploit the indefinte capacity of BV vectors to co-express multiple transgenes and thus modify the glycosylation process in the host by expressing appropriate glycosylation enzymes along with the transgene of interest.
• Example this strategy was used to co-express
fowl plague hemagglutinin and beta-1,2-N-acetylglucosaminyl-transferase resulting in the synthesis of large amounts of hemagglutinin correctly modified with N-acetylglucosamine residues.

• The construction of baculovirus expression vectors involves inserting the transgene downstream of the polyhedron promoter. This is normally achieved by homologous recombination using plasmid vector carrying a baculovirus homology region.
• Baculoviruses can also be used for the delivery of foreign genes to viruses to produce a cell model or animal models for certain diseases. Example the hepatitis C virus does not infect cultured cells, but a hybrid baculovirus containing the entire HCV genome can initiate an HCV infection.

55
Genotyping Transgenic Mice by PCR to Screen for Potential Founders
• This is the test method of mice for the presence of the transgene by PCR.
• Since we know the sequence of the gene that was inserted into the male pronucleus, we could determine if the mouse contains the transgene of interest, by performing PCR.
• Tail biopsies from potentially transgenic mice will be obtained 5 weeks after injecting eggs (3 weeks gestation time and 2 weeks of post-natal growth). The investigator then extracts DNA from the tail tips and tests for the transgenic by PCR. How?

56
• By designing a set of primers that are taken from the transgene sequence and using them in a regular PCR to amplify the gene of interest if found.
• Now if the mouse is a transgenic mouse, then a PCR product corresponding to a known size will appear in the gel. But if the mouse is NOT a transgenic one, there should be NO PCR product corresponding to that size.
• In addition, to evaluate the stability of the insert, some markers has to be checked and the best ones are the ones that can be assayed readily such as observing the new phenotype of the progeny.

57
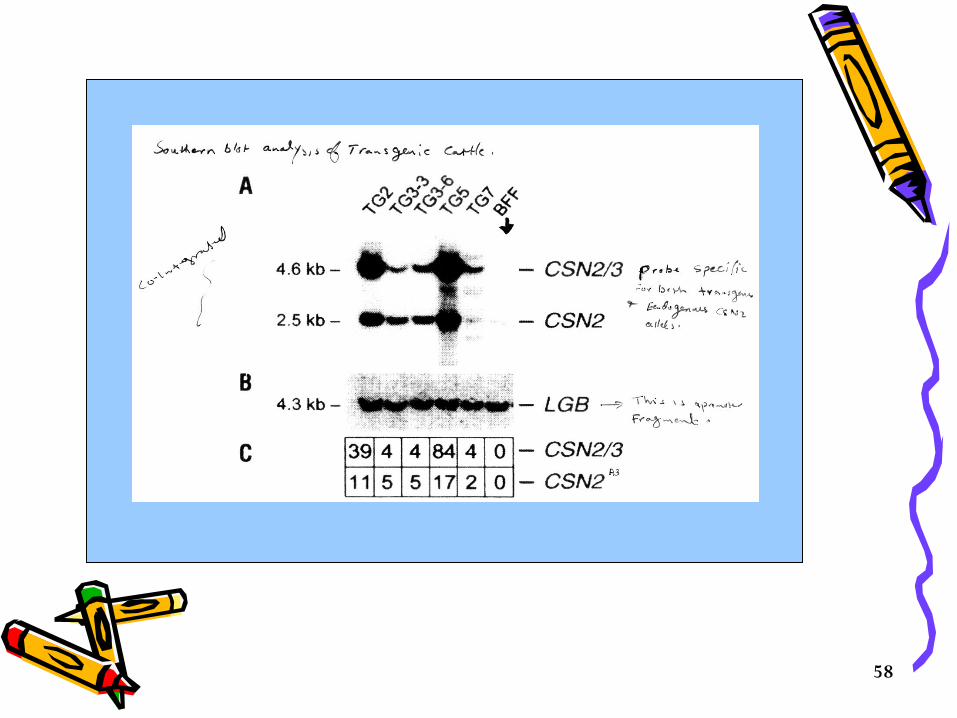
To determine the number of copies and places, Southern Blotting Analysis will be done;
• When pups are 6 weeks old, Southern blot analysis should be done to determine how many copies of the transgene were integrated, how many chromosomal sites the transgene inserted into, to verify transgenic status and to determine if the transgene is intact.
• With this information, transgenic founders with a good chance of transmission (at least 5-10 copies) of an intact transgene in a single insertion site can be selected for intensive breeding. (Figure 1 ).
58

59
• One of the problems is that when DNA is micro-inserted, randomly some parts of it will replace some genes in the mouse, and thus might inactivate them.
• Depends on which gene is inactivated, a damage to the progeny might occur.

60
Engineered Embryonic Stem Cell Method • In this method, cells from the Inner Cell Mass
(ICM) of early embryos blastocysts (a stage of a developing mouse embryo) will be used.
• These cells can be grown in cell culture and still retain the capability of differentiating into other cell types including germ line cells after they are introduced into another blastocyst embryo.
• Such cells are called pluripotent (multi) embryonic stem (ES) cells. These cells can be easily manipulated by genetic engineering without changing their pluripotency.

61
Steps of the procedure;
1. Obtain fertilized eggs (pre-implantation zygotes) from a pregnant mother mouse as described above.
2. Grow zygotes in culture until day 3.
3. Harvest the Inner Cell Mass (ICM) from 3 day old blastocysts.
4. Culture the Inner Cell Mass (ICM) on feeder cells to develop Embryonic Stem (ES) Cell lines.

62
5. Create transgenic ES cells by microinjection or by introducing cells briefly to an electrical potential that disrupts cell membrane thus allows the entrance of DNA containing the transgene that was constructed with the genes of interest.
• In this method a functional transgene can be integrated in the place of a dispensable gene in the genome of the ES cell.

63
6. Inject the transgenic ES cells into the blastocoele (fluid filled cavity of the mass of cells) of a new 3-day old host blastocyst.
• The injected ES cells combine with the host ICM and contribute to the developing embryo.
• The first generation offspring are chimaeras - they have somatic cells composed of both transgenic ES cells and host cells
• And also have germ cells composed of both transgenic ES cells and host cells
• Usually a coat color gene is used in the transgene construct as a visual marker to facilitate the quick detection of the transgenic (chimaeric) pups.

64
7. The transgene, if in the germ cell lineage, can be transmitted to offspring and homozygous transgenic lines can be constructed.
• After transfection of ES cells in culture with the DNA vector;– Some cells will have DNA integrated at none-target
(spurious) sites– Some cells will have DNA integrated at target (correct) sites– Some cells will not have any DNA integration

65
How to enrich DNA integration at the specific sites?
A procedure called positive/negative selection is implemented.
• This procedure used positive selection for cells did not accept the DNA inserts and negative selection for cells who have DNA integrated any where in their genome.
• In this procedure, a construct will be prepared and should contain the following;
– Two blocks of DNA sequences (HB1 and HB2) that are homologous to separate regions of the target site.

66
• The trans gene, TG that will confer a new function on the recipient
• Neor, a DNA sequence that codes for an enzyme that inactivates neomycine and its relatives such the drug G418 which is lethal to mammalian cells

67
• Two different genes for the thymidine kinase (tk1 and tk2). These enzymes phosphorylates the nucleoside analogue called gancyclovir.
DNA polymerase fails to discriminate against
the resulting nucleotide and inserts this nonfunctional nucleotide into freshly-replicating DNA. So gancyclovir kills cells that contain the tk gene.
• Now the arrangement of these sequences is key to the positive and negative selection procedure.

68
Possible results; • Most cells fail to take up the vector; these cells will be
killed if exposed to G418 as the neo gene will not be incorporated. (positive selection).
• In a few cells: the vector is inserted randomly in the
genome. In random insertion, the entire vector, including the tk genes, is inserted into host DNA. These cells are resistant to G418 but killed by gancyclovir. (Negative selection).
• In still fewer cells: homologous recombination by double crossover at target sites occurs. i.e Stretches of DNA sequence in the vector find the homologous sequences in the host genome and the region between these homologous sequences replaces the equivalent region in the host DNA.
• Therefore, tk genes will be excluded and cells survive both G418 and gancylovir as only the neo and the trans genes are included.

69

70
• Now by this method ES cells that carry the target site
will be enriched several thousand fold, thus better chances of producing a transgenic animal with the desirable characters.
• By this method, ES cells that contain the target, are identified and cultured for propagation.
• Embryonic stem cells carrying an integrated transgene can be cultured and inserted into blastocyst stage embryo and these embryos can then be implanted in pseudopregnant foster mother.

71
• Transgenic lines can then be established by first mating founder transgenic mouse to animals from the same strain and then crossing transgenic litter mates to create a homozygous transgenic animal.
• Unfortunately pluripotent ES cells comparable to those of mouse were not found in cattle, sheep, pigs or chickens.

72
Scientific and medical applications of the ES cells method of transfection
• This route has been usually employed to; inactivate a gene, alter it, or replace its protein coding region with a reporter (a coding unit whose product is easily assayed. It may be connected to any promoter of interest so that expression of the gene can be used to assay promoter function).
• Main application of ES cell transgenic mice are to medicine and pure science including;– Improve understanding of all aspects of healthy animal– Understanding therapeutic approaches– Understanding biochemistry and physiology particularly;
mammalian development, neurobiology, learning and memory.
73
Nuclear Transfer Method (non-transgenic method)
• In this process the sheep Dolly was generated from an enucleated (nucleus was removed) egg into which the nucleus from a cultured somatic cell of a mature sheep has been introduced.
Method• Oocytes are recovered from animals between 28-33
hours after injection of gonadotropin releasing hormone,• Oocytes are recovered in PBS containing 1% FCS and
transferred to a new media containing 10% FCS and incubated at 37° C.
• Nucleus is removed manually from an unfertilized oocyte as soon as possible

74
• The somatic cell has to be in a non-dividing stage (G0) why ? ; this can be done in culture by depriving it of external stimuli that provokes growth. How?. Read the provided article
• A non-dividing somatic cell is placed in contact with the oocyte and the two are fused together by applying an electrical potential which also activates the egg thus, mimicking the process of natural fertilization.
• The result of this hybridization is an activated oocyte with two chromosome sets (from the diploid somatic cell).

75
• Usually, the cytoplasm of normal oocyte contains proteins and RNA molecules that are required for the early stages of development but in this case, the cytoplasm of the somatic cell contains a whole set of genes that are reprogrammed to take control over the developmental program in the same way as the genes of the normal embryo.
• Effect of age; it was found that cells obtained from fetuses and new borne donors are more efficient in nuclear transfer while clones derived from adult cells show more abnormalities.
• Why? It could be due to the fact that somatic cells of adult animals have accumulated more mutations or they are more differentiated than fetal cells thus are more likely to fail the full term development.

76
Applications of Nuclear Transfer;
• Nuclear transfer has applications outside the field of transgenesis such as propagation of an animal with a particularly desirable set of genes.
• Since propagating the transgenic animals are
not easy and normally goes with risks, the nuclear transfer can therefore, be used for propagating a successful transgenic animal making a whole herd of that animal !!! ….. Prohibitively expensive.

77
Applications of transgenic animals
Transgenic mice Transgenic mice can be used for;
– As test subjects to determine the effectiveness of potential therapeutic agents
– Although mice are far from humans, some times they can serve as models for human diseases.

78
Specific Applications of Transgenic MiceTransgenic Mice in Oncology
• The study of transferred oncogenes has always been hampered by the fact that cell lines in culture have already been transformed to an abnormal phenotype.
• The ability to insert oncogenes or proto-oncogenes
into embryos and to study their effects in normally differentiating cells of an intact organism has circumvented this problem. Results of such studies have made an enormous contribution to our understanding of neoplastic diseases and its relationship to aberrant gene expression.

79
Transgenic Mice as Animal Models of Human Diseases
Animal models for human illnesses are useful for studying the pathogenesis of diseases as well as for developing and testing new therapies. Human diseases can be induced in transgenic mice by expression of transferred genes, or by insertional disruption of endogenous sequences.
Some examples of models created by transgene expression are listed below.
• Hepatitis B is a human disease that lacks a readily workable animal model. Introduction of the HBSAg gene into mice results in transgenic mice that mimic the carrier state with production of HBsAg in the liver but with an absence of disease

80
Transgenic Mice as Models for Gene Therapy
• Genes can be inserted into transgenic animals and function to alleviate disease states, such model systems can be of great importance in improving our understanding of the potential for gene transfer as an approach to treatment of diseases.
• Mice with growth hormone deficiency are markedly reduced in size and males suffer from infertility. Introduction of the growth-hormone gene into these animals leads to growth which exceeds that of normal animals and restores male fertility.
• However, the pattern of release of growth hormone that results from transgene function is apparently inconsistent with female fertility. i.e does not restore it.

81

82
Another Example• Insertion of either the mouse or human β-globin
gene can reduce the severity of β-thalassemia in mice. In these experiments, the product of the human globin gene was able to associate effectively with the mouse A chains, and it actually functioned better than the transferred mouse gene β-globin in reducing severity the thalassemic state.
• Mice with a deficiency in gonadotropin-releasing hormone (GnRH) are infertile and exhibit profound perturbations of their reproductive endocrine functions. Cloning of the GnRH gene and its transfer into mice has resulted in restoration of normal endocrine function and in fertility.

83
Alzheimer’s disease model
• Alzheimer’s disease is a degenerative brain disorder that is characterized by the progressive loss of both abstract thinking and is accompanied by personality change, language disturbance and a slowing of physical capabilities.
• The brain of those patients accumulate within the body of the neurons a dense material called senile plaques.
• The principal component of senile plaques and amyloid bodies is a 4-kDa protein called βA4 (or β protein). This protein is the product of an internal proteolytic cleavage of the β- amy1oid precursor (APP).
• Researchers have found that some strains of mice produce senile plaques during their life span, whereas others do not. Thus, the later (none producers) strains are important for forming transgenic mice that carry and express a transgene encoding the βA4 portion of APP which might provide a model for studying the molecular basis of Alzheimer’s disease.

84
Importance of such experiment: this type of mice can be used for a precise determination of the mechanism of Alzheimer’s disease and probably for a treatment scheme.
• One of the vectors that has been constructed for modeling Alzheimer’s disease in mice consists of ;– A promoter region from brain specific virus ligated to a
portion of the human APP (β amyloid precursor protein) gene that encodes the last 100 amino acids at the C terminus of APP, which includes the βA4 amino acid sequence.
– Transgenic mice were established with this construct, and expression of the transgene was confined to neurons of the brain.
– Immunocytochemical studies showed that the brain of transgenic mice accumulated βA4 protein that was derived from the transgene. How?

85
• This screening can be done using anti- βA4 antibody that is conjugated to a dye which makes it visible either to the naked eye or under special microscope such as fluorescence.
*******• Alternatively, ES cells that have a site-directed
mutated APP gene could be used to establish a transgenic line that might mimic Alzheimer’s disease more precisely.
• Transgenic mice have also been used as models for expression systems that are designed for secretion of the product of a transgene into milk.

86
Another example (CF)• Another example of the usefulness of the
transgenic mice is the production of large quantities of authentic cystic fibrosis trans-membrane regulator (CFTR) that are needed to study its function and possibly formulate potential therapies for treating cystic fibrosis.
• CFTR normally acts as a chloride channel but when its function gets altered, cystic fibrosis occurs and it will be characterized by the accumulation of mucus into the lungs and pancreas.

87
What is Cystic fibrosis?
• It is the most common lethal human hereditary disorder, occurring once in every 3,000 births.
• It affects the lung, intestinal tract and liver, with thick mucus, chronic airway infections and inflammation beginning in early childhood and leading to progressive loss of lung function.
While the life expectancy of these children is double what it was, they are still only expected to live to 40. The underlying defect is in a gene that codes for a substance that regulates protein secretion across a cell membrane, but infection also plays a major role.
All existing therapies only alleviate the symptoms by reducing infection and mucus.

88
To get large amounts of CFTR; • A full length CFTR cDNA sequence was cloned
into the middle of a defective goat β-casein gene
• The construct retained the promoter and the
termination sequences of the goat β-casein gene.
• The β-casein gene is then actively expressed in mammary glands during lactation producing the β-casein which is the most abundant protein in the milk.

89
• Now, transgenic mouse lines carrying the CFTR sequence under the control of the β-casein gene regulatory sequences were established.
• The product is milk from transgenic females contained the CFTR protein bound to the membrane of fat globules.
• This is a model, however, to obtain mega quantities of
this protein, a construct has to be introduced into a larger animal such as sheep, cows or goat.

90
Antisense Genes in Transgenic Mice
• Another method for negating gene function involves the use of antisense transcripts.
• When genes are cloned in reverse orientation with respect to the promoter, RNA may be produced from the non-coding strand.
• This RNA, presumably by forming a heteroduplex with the sense RNA, can block translation of cytoplasmic mRNA.
• Thus, antisense genes can be used to obliterate (wipe out) production of proteins from specific genes in transgenic animals.
• The feasibility of this approach has recently been demonstrated by the transfer of an antisense construct of the gene for myelin basic protein (MBP) into mice. Interference with the production of MBP resulted in dysmyelination. Although this research is still its infancy, it has great potential for future experiments.

91
Transgenic cattle
• If the mammary gland is to be used as a bioreactor, then dairy cattle are the likely candidates for transgenesis as they produce about 10,000 liters of milk/year with 35 gm protein/liter.

92
Protocol to produce transgenic cattle (Figure 15-9)
• collecting oocytes from slaughterhouse –killed cows• in vitro maturation of these oocytes• in vitro fertilization with bull semen• centrifugation of fertilized eggs to concentrate the yolk so
that male pronuclei will be seen under the dissecting microscope.
• microinjection of input DNA into male pronuclei• in vitro development of embryos• embryo implantation into a recipient foster mother• DNA screening of the offspring for the presence of the
transgene.• When this procedure was done only two transgenic calves
were produced from a starting pool of 2470 oocytes which means that the procedure is feasible but in efficient in this format.

93
Goals of producing transgenic cattle
• To change the constituents of milk. For example the amount of cheese produced from milk is directly proportional to the amount of k-casein content of the milk so if a transgene is constructed to produce milk with higher amounts of k-casein, then the production of cheese will increase proportionally.
• Production of transgenic cows with modified genes to produce lactose free milk could solve the problem of those who have lactose intolerance.
• For livestock in general, attempts to produce animals with inherited resistance to bacterial, viral, and parasitic disease is a goal. Example of major diseases that affect the livestock are mastitis in cows, neonatal dysentery in swine, fowl cholera.
• If the basis of each of these is a single gene that will be
responsible for the resistance, then it might be possible to produce transgenic animals that carry this gene.

94
Other alternative, is the production a transgenic animal with inherited immunological protection.
A number of candidate genes that contribute to the immune system such as Major histocompatibility genes, T-cell receptor genes, lymphokine genes are under study to evaluate this potential.
But the most favorable preliminary results to date comes from research in which the genes encoding the heavy and light chains of a monoclonal antibody (MAb) have been transferred to mice, rabbits and pigs.

95
• By this introduction of MAb, these animals will have an endogenous source of MAbs with predefined specificity toward certain pathogen, thus eliminates the need for immunization. This concept is called In vivo immunization.

96
• Example; the genes for the immunoglobulin chains of a mouse MAb that are specific to “ 4-hydroxy-3-nitrophenylacetate were cloned in a tandem and microinjected into fertilized egg of mice, rabbits and pigs.
• In each case MAb activity was found in the serum but the concentrations of the antibodies were low which could be due inheritable problems of the construct, thus a new construct should be tested.

97
Transgenic Sheep• Transgenesis research with sheep, goat or pigs
has concentrated in the most part on utilizing their mammary glands as bioreactors for production of pharmaceutical proteins.
Example; Production of transgenic sheep that produces anti-trypsin in their milk; This protein is a potential treatment for cystic fibrosis.
The Technology• PPL Therapeutics transfers genetic material from
one organism to another using the same technology it used to produce "Dolly the Sheep", a process called somatic cell nuclear transfer.

98
Steps of the procedureGenes are Modified: A single cell from a sheep is
modified to include the human gene for the protein alpha-1 antitrypsin. However, the gene must only turn on in the mammary glands so that the protein only appears in the sheep's milk.
• Before the sheep DNA is modified, the human gene is fused to the promoter gene for beta-lactoglobulin. The human gene will only be expressed when the beta-lactoglobulin is turned on, and this only happens in the milk-producing mammary glands.
Injection : The nucleus, containing the modified DNA, is removed from this cell and injected into the enucleated fertilized sheep oocyte. Or the modified somatic cell could be fused to the enucleated oocyte.

99
Implantation of embryos: The fertilized sheep embryo is implanted into a surrogate mother for the rest of its pregnancy.
Lactation: Upon giving birth to a lamb the mothers (ewes) produce milk (lactated). Beta lactoglobulin production started during lactation, so did production of human alpha-1 antitrypsin. The rams also contain the required gene but it is not active, although it can be passed to their offspring. The alpha-1 antitrypsin protein that is expressed in the milk can be extracted and purified.
Next Generation: The newborn lambs were screened for presence of the gene (by DNA analysis of tail tissue or blood from the jugular) and mated when mature.
The production flock was started from semen from two transgenic rams brought to New Zealand in 1996. Conventional New Zealand ewes were inseminated and some of the resulting lambs were transgenic. Embryos from transgenic animals were transferred to surrogate mothers.

100
The modified gene is shown to be stable (i.e has been transmitted faithfully from parent to offspring). How can we judge that?
• Homozygotes are as healthy as heterozygotes; this shows that the gene has not inserted into an essential part of their genetic material - insertion into other parts of the DNA would lead to death of the offspring
• The human protein secreted in the milk has been consistent in quantity and quality

101
The Benefits• The most obvious benefit from this
research is:• Production of a treatment for cystic
fibrosis. How? See next slide

102
Human alpha-1 antitrypsin is currently derived from blood plasma and administered intravenously at 60mg/kg once a week. The difficulties with this treatment are:
1. Cost - treatment for an individual costs $40,000 per year2. Availability - the protein is produced in plasma at a concentration
of about 1.5 g/L and obtained from healthy donors.3. Contamination - any extraction of material from blood carries
risk of contamination from other diseases such as HIV, new variant Creuzfeldt-Jakob Disease (BSE) and Hepatitis B.
4. Efficiency - using transgenic animals produces far greater quantities of the protein at lower cost in the long term, this research will also provide further benefits:• Provide techniques for producing other disease-fighting drugs• Provide techniques for incorporating medicines in foods• Help scientists to understand how milk protein is produced and modified Transgenic-derived proteins were glycosylated and had biological
activities comparable to those extracted from human sources.

103
Goats and PigsGenerally the production of transgenic goats and
pigs is similar to that for sheep however, there are some differences in that the; – expression of transgenes in the mammary glands of
sheep or goats had no ill effects on either lactating female or nursing progeny.
– While the transgene for bovine growth hormone-under the control of the metallothionine promoter- when introduced into pigs, several adverse results were observed;
• Gastric ulceration• Kidney dysfunction• Lameness• Inflammation of the lining of the heart• Swelling of the joints • Susceptibility to pneumonia

104
Transgenic Birds• Avian ova are normally fertilized approximately
30 minutes after ovulation. Cell division occurs in the oviduct for approximately 20 hours before ovi position. At this time, the embryo is comprised of approximately 60,000 pluripotent cells, which are collectively called the blastoderm.
• The presence of a large yolk and multiple
pronuclei makes direct microinjection of DNA impractical.
• Therefore, DNA microinjection into fertilized bird eggs to produce transgenic strains is not possible;

105
• During fertilization in birds several sperms can penetrate the ovum, instead of only one as in case of mammals.
• Therefore, it is not possible to identify the male pronucleus that will fuse with the female pronucleus.
• Microinjection of DNA into cytoplasm is not enough for
the process to proceed as the DNA will not integrate into the genome of the fertilized egg.
• The technique also would be difficult as the avian ovum after fertilization become enveloped in tough membrane and surrounded by large quantities of albumin and enclosed in inner and out shell membranes.

106
• By the time the avian egg outer shell membrane hardened, the developing embryo (blastoderm stage) will be two layers of 40,000 to 80,000 cells.
• At the moment no one has identified avian specific embryonic stem cells so this approach can not be used in birds. The alternative is a procedure using engineered cells from embryos. How??

107
ProcedurePlastoderm cells are removed from the donor chickenThese cells get transfected with cationic lipid (liposome) transgene DNA complexes (lipofection).The cells will be reintroduced into the subgerminal space of embryos of freshly laid eggs. Figure 15-10 shows a schematic diagram of this procedure.
Some of the progeny will consist of a mixture of cells with some cells from the donor but most from the recipient, such mixture is called the chimera
* Lipofection: delivery into eukaryotic cells of DNA and RNA or other compounds that have been encapsulated in an artificial phospholipid vesicle.

108
Now in some of these chimeras cells that were descended from transfected cells may become part of the germ line tissue and form germ cells.Transgenic lines can then be established by rounds of mating.The proportion of chimeras can be increased to enhance the probability of obtaining germ line chimeras if the receiving embryos are irradiated with a dose of 540-660 rads for 1 h prior to the introduction of transfected cells. Irradiation destroys some of the blasoderm cells thus increasing the final ratio of the transfected cells to the recipient cells.

109
What can we use transgenic chicken for?To improve the genetic makeup of the existing strains with respect to
resistance to avian viral and coccidial diseases, better feed efficiency, lower fat and cholesterol in eggs and better meat quality.
The egg with its high protein content could be used as a source of pharmaceutical proteins

110
Transgenic FishAs natural fisheries become exhausted, production of this source will depend more on the aquaculture. Production of transgenic fish therefore become a primary objective.
To date, transgenes have been introduced by DNA microinjection into the fertilized eggs of number of fish species including; Catfish , Crap, Trout , Salmon , Tilapia In fish the pronuclei are not readily seen under the microscope after fertilization, therefore, a linearized transgene DNA is microinjected into the cytoplasm of either fertilized eggs or embryos that have reached the 4 cell stage.

111
Now because fish eggs develop externally there will be no need for implantation. Instead the development can be done in the Temperature regulated tanks with a survival rate from 35-80% and production of the transgenic fish ranges from 10-70%. Same as in transgenic animals, the founder fish can be mated and transgenic lines established.
In one study, a transgene consisting of the promoter region of the antifreeze protein gene of the fish called ocean pout. The growth hormone cDNA from salmon. And the termination polyadenylation signals from the 3’ end of the end of the antifreeze protein. This construct was injected into eggs of Atlantic salmon.

112
Result; the transgenic salmon was larger and grow faster than the none transgenic.Eventually, genes for disease resistant, tolerance to environmental stress, and other biological features will be introduced into fish in cold and warm waters.

113
Thank you…Do you think you
have learned some thing new?


















