
1 9 7 7 - Open Repositoryarizona.openrepository.com/arizona/bitstream/10150/348230/1/AZU_TD... ·...
58
Effect of temperature and photoperiod on the reproductive biology and form determination of the blue alfalfa aphid, Acyrthosiphon kondoi Shinji Item Type text; Thesis-Reproduction (electronic) Authors Kodet, Russel Todd, 1952- Publisher The University of Arizona. Rights Copyright © is held by the author. Digital access to this material is made possible by the University Libraries, University of Arizona. Further transmission, reproduction or presentation (such as public display or performance) of protected items is prohibited except with permission of the author. Download date 28/08/2018 10:55:39 Link to Item http://hdl.handle.net/10150/348230
Transcript of 1 9 7 7 - Open Repositoryarizona.openrepository.com/arizona/bitstream/10150/348230/1/AZU_TD... ·...
Effect of temperature and photoperiod on thereproductive biology and form determination of
the blue alfalfa aphid, Acyrthosiphon kondoi Shinji
Item Type text; Thesis-Reproduction (electronic)
Authors Kodet, Russel Todd, 1952-
Publisher The University of Arizona.
Rights Copyright © is held by the author. Digital access to this materialis made possible by the University Libraries, University of Arizona.Further transmission, reproduction or presentation (such aspublic display or performance) of protected items is prohibitedexcept with permission of the author.
Download date 28/08/2018 10:55:39
Link to Item http://hdl.handle.net/10150/348230

EFFECT OF TEMPERATURE AND PHOTOPERIOD ON THE . REPRODUCTIVE BIOLOGY AND FORM DETERMINATION
OF THE BLUE ALFALFA APHID, ACYRTHOSIPHON KONDOI SHINJI
byRussel Todd Kodet
A Thesis Submitted to the Faculty of theDEPARTMENT OF ENTOMOLOGY
In Partial Fulfillment of the Requirements For the Degree ofMASTER OF SCIENCE
In the Graduate CollegeTHE UNIVERSITY OF ARIZONA
1 9 7 7

STATEMENT BY AUTHOR
This thesis has been submitted in partial fulfillment of requirements for an advanced degree at The University of Arizona and is deposited in the University Library to be made available to borrowers under the rules of the Libraryo
Brief quotations from this thesis are allowable without special permission<, provided that accurate acknowledgment of source is made. Requests for permission for extended quotation from or reproduction of this manuscript in whole or in part may be granted by the head of the major department or the Dean of the Graduate College when in his judgment the proposed use of the material is in the interests of scholarshipo In all other instances3 however3 permission must be obtained from the author.
APPROVAL BY THESIS DIRECTOR This thesis has been approved on the date shown below:
"M. Wo NieLbon Professor of Entomology
Date

ACKNOWLEDGEMENTS
I wish to sincerely thank Dr. M. W» Nielson of the U. So Do Ao for his constant help in the. planning of this study and criticism in the preparation of this manuscripte His guidance is gratefully acknowledged„.
I would also like to thank Drs. R. E. Golds R„ To Huber5 and Go W. Ware for their suggestions and for serving on my graduate committee« I appreciate the help of Dr.Ro OoKuehl in analyzing these datao
Sincere thanks are also extended to; my wife, Linda = My graduate study would not have been possible without her constant encouragement 3 support, and help«
iii

TABLE OF CONTENTS
PageIjXiSl? OF TASIjES o o o o o o o o . o „ o O O o o o o o o . o o ViLIST OF ILLUSTRATIONS o o * * * * * * „ «, * „ , * o * viiABSTRACT o © © © © © © © © © © © © © © © © © © ' © © © © y**! xt.. INTROJ3U0TI ON © © © © © © © © © © © © © © © © © © © © © ILITERATURE REVIEW © o © © © © © © © © © © © © © © ©'© 2
Rl St/ O^y © © O o © o o © o o © o o o o o © o o © o 2Economic Importance © © © © © © © © © © © © © © © 2Taxonomy © © © © © © © © © © © © © © © © © © © © 3.Reproductive Biology . o •«;«;«. . • « ........ 4
Temperature <> o.--. <>_ o o. o, ot .o © © © © © ©, ©. © © 4Photoperxod © © © © © © © © © © © © © © © © © ^
Form Determination © © « © © © . « © © © © . © © 7Temperature © © © © © © © © © © © © © © © © © 7Photoperxod ©© © © ©© © © © © » © © © © © © 9
MATERIALS AND METHODS © » © © » © © © © © © © © © . © 10Environmental Chambers . . © © „ © . . © = . . » . 10Host Plants » » . « » » » © © . . . . © « © . . . © 11"CageS © 6 © © o © o o © o e © © o o © © o o e © o 11Temperature/Photoperiod Treatments © . « . . » © » 12Experimental Aphids . . © . . © . « . » = » „ » « 13Statistical Methods . „ © © © « , „ © © » © © . . 14
RESULTS AND DISCUSSION © © . . . © © , © . . . » © . . 15Number of Instars » © « © » © , © © « » » . , . » 15Length of Nymphal Development . © » © „ © , « . » 15Nymphal Survivorship 21Preparturition Period © © © © © © © © © © © © © © 23Parturition Period , © © © , . © , „ . © © © © © » « 23Postparturition Period » © . © „ © . . . . . . . , 25Adult Survivorship » . © „ » © © . » . © « » . » = 25
© © © © © © © © © © © © © © © © © © © © "7Intrinsic Rate of Increase « » „ , © » » » » . « , 30Doubling Time © © 32Form Determination © » » © » © © © » « © « © = © = 32
iv

VTABLE OF CONTENTS--Continued
PageCONGLNSXONS «•.. •... •«- • «... «... o/ o■ «. « » • .• », « o ». •«... «> ®. • 40SUMMARY o o o o o o o o o' o o o o o o o o o o o o o o ' .
LITERATURE CITED « « ® o o o «. ® ® o o ® » o » ® « ■ 44

LIST OF TABLES
Tablelo
20
3.
5.
60
?<>
80
9-
10.
PageThe Effect of Temperature and Photoperiod
on Nymphal Development of A. kondol . 0 . » ... 16The Effect of Temperature and Photoperiod
on the Mean Number of Day-Degrees Requiredfor A. kondoi to Complete Nymphal■Development” 18
The Effect of Temperature and Photoperiod on the Mean Duration of Four Instars of A-o kondoi . . . . . . © © © © © © © © © © 20
The Effect of Temperature on NymphalSurvivorship of A© kondoi . © © © © © © © © © 22
The Effect of Photoperiod on NymphalSurvivorship of A. kondoi © © © © © © © © © © © 22
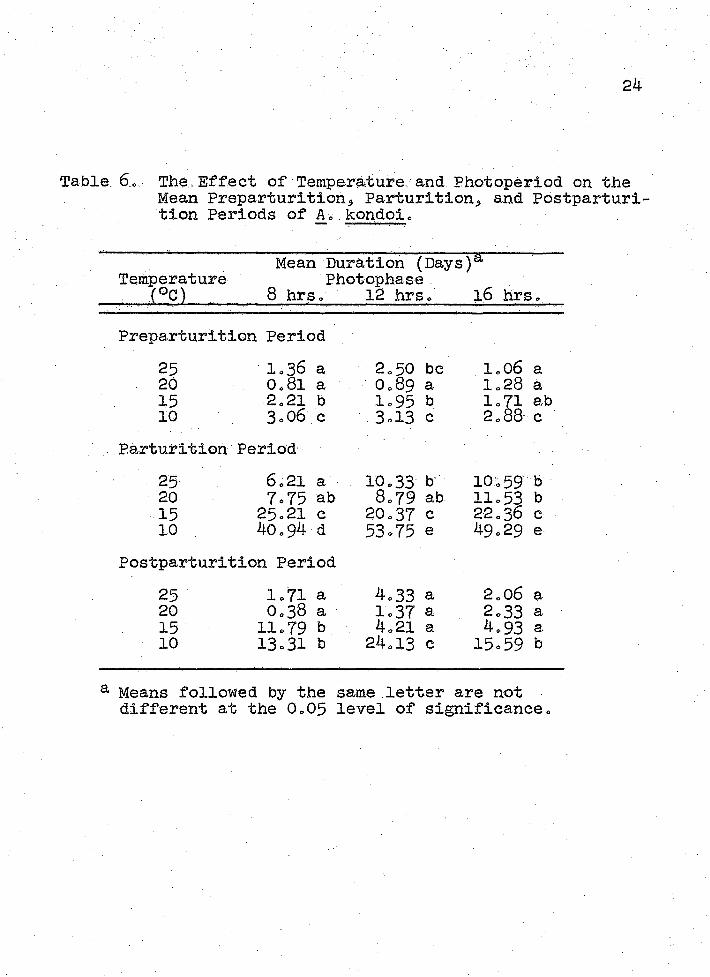
The Effect of Temperature and Photoperiod on the Mean Preparturitions Parturition, and Postparturition Periods of A© kondoi © © 24
The Effect of Temperature and Photoperiod on the Mean Fecundity per Female and the Mean Fecundity per Female per Day o d? .A © kondoi © © © © © © © © © © © © © © © © 28
The Effect of Temperature and Photoperiodon the Doubling Time of A© kondoi „ © © © © 33
The Effect of Temperature and Photoperiod on the Length of the Reproductive Period of A© kondoi and on the Time at which Form Reversal Occurred © © © © © © © © 37
Reproductive Age at which 70$ of the TotalReproduction of A, kondoi Had Occurred © „ © 39
vi

LIST OF ILLUSTRATIONS
Figurel.o The Effect of Temperature and Photoperiod
on Adult Survival of A„ kondoi2. The Effect of Temperature and Photoperiod
on the Intrinsic Rate of Increase of Ao kondoi O O O O O O O 0 , 0 o o o o o o <
The Effect of Temperature and Photoperiod on the Predominant Alary Form Produced Early in the Reproductive Period of Ao kondoi O O O €> » 6 o o o o o o o d o <
The Effect of Temperature and Photoperiod on the Predominant Alary Form Produced Late in the Reproductive Period.of A. kondoi e o o o o e o o o o o o o o o <
vii
Page
, o 26
o o 31
o . 35
o o 36

ABSTRACT
The biology and form determination of the blue alfalfa aphid3 Acyrthosiphon kondoi Shinji (Homoptera; Aphid- idae)a an important pest of alfalfa^ Medicago sativa L ea were investigated at 5 constant temperatures and 3 photoperiods on Melilotus officionalis Lam..
The length of the nymphal and adult stages decreased as temperature increased from 10° to 250C. Nymphal survival was greatest at 20°C and 12 hours photophase3 while no survival occurred at 30°G. Total fecundity increased as temperatures decreased from 25° to 10oCs but the fecundity rate increased as temperatures increased from 10° to 25°C. The optimum conditions for populations to increase occurred at 25°/l6 hours photophase. Photoperiod affected all nymphal and adult parameters at low temperatures s whereas photoperiod had little or no effect on the biology of A. kondoi at high temperatures.
No sexual forms were produced. Apterous partheno- genie aphids were produced at high temperatures while alatae were produced at low temperatures. Adults switched from the production of one alary form early in the reproductive period to the production of the opposite form later in the reproductive period.
viii

INTRODUCTION
The blue alfalfa aphid (Acyrthosiphon fcendoi Shinji)3 commoriiy found from. Japan through Mongolia on legume speeies of Medicago9 Trifolium9 and Melilotuss was introduced into California in 1974. During 1975 the aphid spread rapidly from California to Arizona and Nevada where it caused severe damage to many alfalfa fields. In 1976 the aphid was reported in Utah3 New Mexico3 and Kansass and by 1977 it had spread into Oregon3 Oklahoma and Nebraska.
The biology of A. kondoi is unknown. Studies were done in the laboratory at Tucson3 Arizona3 to define A. kondoi developmental and reproductive parameters so that population dynamics and polymorphism of the species might be better understood.' In the present study the effect of controlled temperatures and photoperiods were observed on the reproductive biology and form determination of the aphid. These data are presented herein as a predictive model of the response of field populations to these parameters.
1

LITERATURE REVIEW
HistoryAcyrthosiphon kondoi was first reported; in the Impe
rial Valley of California in March* 1975 (Sharma* Stem* and Hagemann 1975)» Examination of preserved insect samples from alfalfa collected in 1974 near Bakersfield* California^ showed the presence of mixed populations of A. kondoi and the pea aphid* Acyrthosiphon pisum (Harris)o However* collections of insects taken from alfalfa in the Imperial Valley during the spring of 1973 contained only A, pisum (Kono1975l Sharma* Stern* and Hagemann 1975; Cooperative Econom-> ■ • 1 1ic Insect Report 1975)» Thus* it appears that 1974 is the earliest year on record of A. kondoi in the United States=
Economic Importance Plants damaged by A. kondoi showed severe stunting
and marked leaf chlorosis within two weeks after initial aphid infestationo Recovery and regrowth of alfalfa was retarded about 50$ during the first two weeks after A. kondoi were killed (Hielson and Kodet 1975; Nielson* Moore* and Kodet 1976). Acyrthosiphon kondoi occurred in damaging numbers in early spring on rapidly growing alfalfa terminals. Populations declined at the onset of warm spring weather. Build-up of damaging populations during fall months has

not been observed (Cooperative Economic Insect Report 1975I Cooperative Plant Pest Report 19765 1977)=
TaxonomyShlnji and EbAdo (1938) described kondol from
Manchuria5 but their co-types were destroyed in World War IX bombingso Tao (1963)5 who was unfamiliar with Shinji's co-typess suppressed A. kondol as a synonym of Aulacofthum solani (Kaltenbech)= Takahashi (1965)5 who was familiar with the destroyed co-types (Eastop 1971s Dickson 1975)s re- described A. kondoi using aphids from Medlcago satiya Lo and Trifolium species„ Acyrthosiphon kondoi was stabilized in two revisions since 1965 (Eastop 1971s Miyassaki 1971)„ Confusion developed over the taxonomy of the species when the aphid was introduced into the United States <, The aphid was identified on separate occasions as Acyrthosiphon sp. -loti groups Ao caraganae (Cholodkovsky)5 and A 0 brevlcaudatus Takahashi before British homopterist V. F. Eastop confirmed the identification of the aphid as A. kondoi. California entomologists proposed the name blue alfalfa aphid9 although the name has not yet been accepted as the common name for the speciesc

Reproductive BiologyAlthough the biology of A„ kondoi is unknowna a
review'of the literature of a, closely related speciess A» pisum, is being presented to establish a basis for comparing and differentiating these common pests of alfalfa*
TemperatureDevelopment and reproduction of A. pisum occurred
within a limited temperature range» No reproduction and development was observed above 30°C (Kenten 1955) or below 5gG (Campbell et al» 1974). Slow reproduction, slow development and high mortality occurred slightly above the lower threshold temperature» Increased reproduction, increased rates of development, decreased mortality, and decreasing longevity were observed with increasing temperatures (Kilian and Nielson 1971)« Increasing temperatures past 20°C (Kenten 1955) or 24»4°C (Kilian and Nielson 1971) continued to decrease longevity, which in turn decreased total fecundity despite increasing rates of reproduction. Fecundity and developmental rates decreased near the upper threshold temperature (Kenten 1955)«
Therioaphis maculata (Buckton) (Messenger 1964) and A. pisum (Siddiqui, Barlow and Randolph 1973) reared at constant temperatures had lower growth, survival, and fecundity than did aphids reared in rhythmically fluctuating temperatures.

Insect developmental rates can be quantified by thermal constants which are the number of day -degrees (heat units) required for individuals of one species to complete development (Gaylor and Sterling .1975)° Day-degrees are found from the equation K = (T - t)d| where,; K is the number of day-degrees5 T is the temperature at which individuals are reared, t is the threshold temperature of development5 and d is the number of days required to complete development » The developmental threshold is found by plotting the regression line for the rate of development (I/days) versus temperatureo The developmental temperature threshold is defined as the point where the regression line intercepts the temperature axis. Acyrthosiphon pisum from Berkeley3 California required 105 day-degrees (t - 5>1°G) to complete development; from Kamloops5 British Colombias 104 day— degrees (t = 5°6°C) and from Vancouver3 British Colombia 118 day-degrees (t - 4„0°C) were required (Campbell et al,1974),
Birch (1948) proposed the intrinsic rate of increase (rm ) as the statistic which best describes optimum conditions for population increase. The intrinsic rate of increase is defined as the exponential growth rate of a population which has a stable age distribution, is living under specified environmental conditions«, and has unlimited space in which to grow. The rate is expressed by the

integrated formula 5% = N0erni*; where: N0 is the number of individuals in time zero, is the number of individuals in time ts and e is the base of natural logarithms. An accurate estimate of rm is obtained from the equation
ix mx dx = 1; where; lx is the probability of a female being alive at age x5 and mx is the mean number of offspring produced in a unit of time of a female aged x. The value of rffl is more easily estimated from the formula ^ e " rmx ixiax = 1, Values for lx and mx are obtained from age specific schedules of births and deaths,
The doubling time is the length of time required for a population to double in number when the population has a stable age distribution^, environmental conditions are constant^ and Space is unlimited (Birch 1948)« The doubling time is found from the equation 0 ,69315/ ^ = t; where; rra is the intrinsic rate of increase and t is the time required for the population to double in size.
PhotoperiodPhotoperiod affected developmental rates5 population
growths and fecundity of A. pisum. The pea aphid produced fewer progeny at high temperatures (30°G) as photophase increased (Kenten 1955)? while at intermediate temperatures (15-20°C) A, pisum produced fewer progeny as photophase decreased (Murdie 1969as 1969b; Sharma5 Larrivee and Theriault 1973s Elliot and McDonald 1976), Murdie (1969a) correlated

■ , . . . • 7decreased fecundity of A. plsum with decreasing body size.He also (1969b) correlated short photophase with significantly decreased body size» Therefore, short photophases decreased fecundity of A. plsum* Elliot and McDonald (1976) found that short photophases promoted alafe development, produced smaller aphids, and reduced fecundity„
Elliot and McDonald (1976) reported alatae developed more slowly than apterae« Accordingly, short photophases which promoted alate development reduced developmental rates» Kenten (1955) also found developmental rates at low temperatures (11°C) decreased as photophase decreased*
Form Determination A wide variety of factors influenced sexual/vivip-
arous polymorphism and alary polymorphism of a number of aphid species (Lees 1966, Hille His Lambers 1966), These factors were crowding, nutrition, plant sap ionic strength, ant relationships, flight, plant age, adult size, adult form, temperature, and photoperiod*
TemperatureTemperature had no effect on alate production in
Aphis gossypii Glover (Reinhard 1927) or A. chlorls Koch (Wilson 1938)o However, high temperatures suppressed alate production in several aphid species: Acyrthosiphon pisum(Harris) (Gregory 1917s Kenten 1955)s Aphis craccifora Koch

8(Johnson 1966)5 Brevlcoryne brassicae L . (Lamb and White 1966)5 Ghaetosiphon fragaefolii (Cockerell) (Schaefers and Judge 1971)a Hyalopterus prunl (Geoffry) (Smith 1937)s Macro- siphum euphoribacae (Thomas) (Shull 1942s MacGillivary and Anderson 1964) 3 sanborni (Gillette) (White 19463 Kitz-railler 1950)3 Megoura viciae Buckton (Lees 1959s 19635 1967)5 Rhopalosiphum madis (Fitch) (Gall 1918)3 and R. padi L. (Dickson and Glen 1971)« Intermediate temperatures lowered the percentage of alatae of Rhopalosiphum fitchii (Sanderson) (Ewing 19163 Wadley 1923s Ackerman 1926).
Low temperatures favored the production of sexual forms in those aphid species which had sexual forms (Lees 1959s Sharmas Larrivee and Theriault 1973) Kenten 1955; Lamb and Pointing 1975)» The temperature effect was greatly enhanced by short photophases. Megoura viciae required a shortened critical photophase of O.25 hours for each 5°C rise in temperature to produce sexual forms (Lees 1964)5 whereas Rhopalosiphum padi L. required 1.75 hours less light for every 4°C rise in temperature to continue production of sexual forms (Dickson and Glen 1971)® Temperature acted directly upon early nymphal stages (Lees 19663 Dickson and Glen 1971) or on the embryo (Hille His Lambers 1966) to affect a humoral regulator.

■' ' 9Photoperiod
The first report of an effect of photoperiod on insects was by Marcovitch (1924) when he showed that sexual forms of Aphis forbesi Weed were produced in response to short daylengths. Similar results were reported for aphids with or without host plant alternation: Megoura viciaeBuckton (Lees 1963)5 Acyrthosiphon pisum (Harris) (Kenten 19551 Sharmas Larrivee and Theriault 1973I Lamb and Pointing1975)5 and Rhppaloslphum padi L, (Dickson and Glenn 1971)«
Lees (1964) located the photoperiodic receptors in the posterior part of the head, of Megoura viciae by the application of strongly localized,light» Photoperiod was perceived both prenatally by an embryo and postnatally by the nymph and adult through the body wall of the female.It has not been shown that the alate/apterae photoreceptor is the same as the sexual/parthenogenetic photoreceptor.

MATERIALS AND METHODS
Environmental Chambers Experiments were conducted in four Percival E-5#U
environmental chambers» Chambers were divided into two levels by a wire rack. Cage height was approximately 15 cm below the light source for the upper levels and- 60 cm for the lower level„
Light was supplied by eight cool-white 36 inch long 50 watt fluorescent tubes and four 75 watt incandescent bulbs* Lights were*separated from-the chamber by a*thermal barrier of translucent plastic which diffused light and isolated lamp heat from the chamber area. Lamp heat caused + l-2°C temperature fluctuations between photophase and scotophase. Light intensity averaged ll6o foot candles 15 cm from the barrier (range 98O-130O foot candles) and 247 foot candles 60 cm from the barrier (range 210-300 foot candles). Adult studies (preparturitions longevity5 fecundity and form determination) were conducted on the upper level5 while nymphal studies were done on the lower level.
Relative humidity was raised by placing trays of water in the 2003 25° and 30°C chambers to approximate the humidity of the 10° and 15°C chambers. Relative humidity
10

11ranged between 60-100$ during each 24 hours in each chamber„ Humidity fluctuations were caused by lamp heat.
Host PlantsPreliminary studies showed that Ap kondoi’preferred
several host plant species over alfalfa0 One of these3 Melilotus offlcionalis (L,) Lam,3 was selected for experimental use due to aphid preference^ favorable growth habit 3 and adequate growth in the environmental chambers„
Host plants were propagated from a single plant, Hooted cuttings were transferred to l4 cm diameter plastic pots containing equal parts of earth3 sand and sphangnum peat moss and were reared in the greenhouse under a photophase of 16 hours and temperatures ranging from 10° to 30oC, Plants were used when four lateral shoots had developed over 10 cm in length. Plants were discarded after each experimental use,
CagesIndividual aphids were confined on terminal stems
in 7,5 cm long #36 dialyzer tubing cages. Developing side shoots were first trimmed off below the terminal leaf bud and then the stem was inserted into the cage. Foam plugs inserted around the stem sealed the cages to prevent aphid escape. New developing side shoots were trimmed away as stems elongated. The entire cage was moved along as the

12stem elongated to maintain the same relative position from the terminal bud. Apical buds were not used since first instar nymphs were difficult to locate in the dense cluster of leaves forming the bud. Four cages were used for each plant.
Cages were transferred to new stems when old stems were broken by handling or damaged by aphid feeding. Such transfers were infrequent and were required only at 10°C where aphids lived and reproduced for long periods of time.
Temperature/Pfaotoperiod Treatments Fifteen combinations of temperature and photoperiod
were selected to explore the biology of the aphid. Five temperature regimens (10o5 15°a 20°s 25° and 30°C)s and three photophases (8S 12 and 16 hours per 24 hour period) were used. The order in which treatments were run was chosen at random. Treatments 10°/12 hours5 20°/l6 hours5 and 25°/12 hours were repeated owing to mechanical failuress logistic problems a and mite outbreaks
Data were collected every 24 hours for all treatments except the reproductive data at 10°C which were taken every 48 hours during the last half of the reproductive period.All other parameters at 10°C were read at 24 hour intervals. At 10°Ca aphids were placed on new plants after reproduction had terminated.

Experimental Aphids A population of A, kondoi was obtained from Reno,
Nevada in June, 1975 and caged in greenhouses in Tucson, Arizona«, Offspring from seven individuals identified by Vo Fo Eastop as A» kondoi were used to establish cultures on Medicago sativa L« and Melilotus offIclonalis»
Aphids were pre-conditioned to all temperature/photo phase treatments in the chambers on M, officionalis prior to data collectiono Several adults were placed in one dialyzer tube cage to begin each treatment» Twenty-four.hours later, one-day old nymphs were transferred to individual cages to start the first. generation. Offspring of the first generation were transferred to 20 individual cages to start a second generation. Nymphal development and survival data were collected from the third generation.
Twelve offspring from second generation aphids were caged in groups of three on terminal buds for form determine tion studies. Offspring from second generation aphids were caged according to how many days the adult had been reproducing. Nymphs were saved on day 1, 4, 7, 10, 13s 16 and 19 of reproduction to follow any change in forms produced over the reproductive life of the female. No nymphs were saved beyond day 19 due to space limitations in environmental chambers.

Statistical MethodsMultivariance analysis was performed by computer
using Multiyariance Analysis Version 5 of the copyrighted program of National Educational Resources, Inc., 215 Kenwood Aye., Ann Arbor, Michigan. Five replications were performed per treatment, and each plant constituted a replication in the analysis. The F-test was used to determine if significant differences existed at the 0.05 level within a treatment parameter, and Least Significant Difference values at the 0.05 level were used to identify those differences(Snedecor and Cochran 1967)«
The Chi Square test at the 0.05 level of significance was used to analyze nymphal survivorship data. The intrinsic rate of increase was integrated by computer from the
X eoe-rmx lxmx , as previously defined.
When the temperature/photoperiod interaction effect was insignificant, data were analyzed to assess the effect of each environmental factor alone. This was done by summing the observations at each temperature across all three photophases, and conversely, by summing the observations at each photophase across all four temperatures. The subsequent results were then subjected to analysis of variance.

RESULTS AND DISCUSSION
Number of Instars Acyrthosiphon kondoi nymphs required four instars
to complete development at all treatments5 except at 30° and 25°C/12 hours photophase where 2 of the 13 nymphs required five instars to complete development» No development or survival occurred at 30°C. The supernumary moults which occurred at 25°/l2 hours photophase indicated that the normal biology of A. kondoi had been disrupted and that the upper threshold temperature was being approached
Whereas Ao pisum did not expedience supernumary moults at 25°C5 it appears that A» kondoi development would be less successful at high temperatures than would be that of A. pisum. Accordinglys in a pest management situations A. pisum would develop more successfully than would A, kondoi during the warmer months of the alfalfa growing season.
Length of Nymphal Development The length of nymphal development was optimum at
25°/l8 hours and slowest at 10°/8 hours photophase (Table 1)« Nymphal development was slower at 10° and 15°C at either 8 or l6 hours photophase« Converselys at 20° nymphal development was significantly faster at either 8 or l6 hour photo- phases than at 12 hours photophase. The length of the
15
16
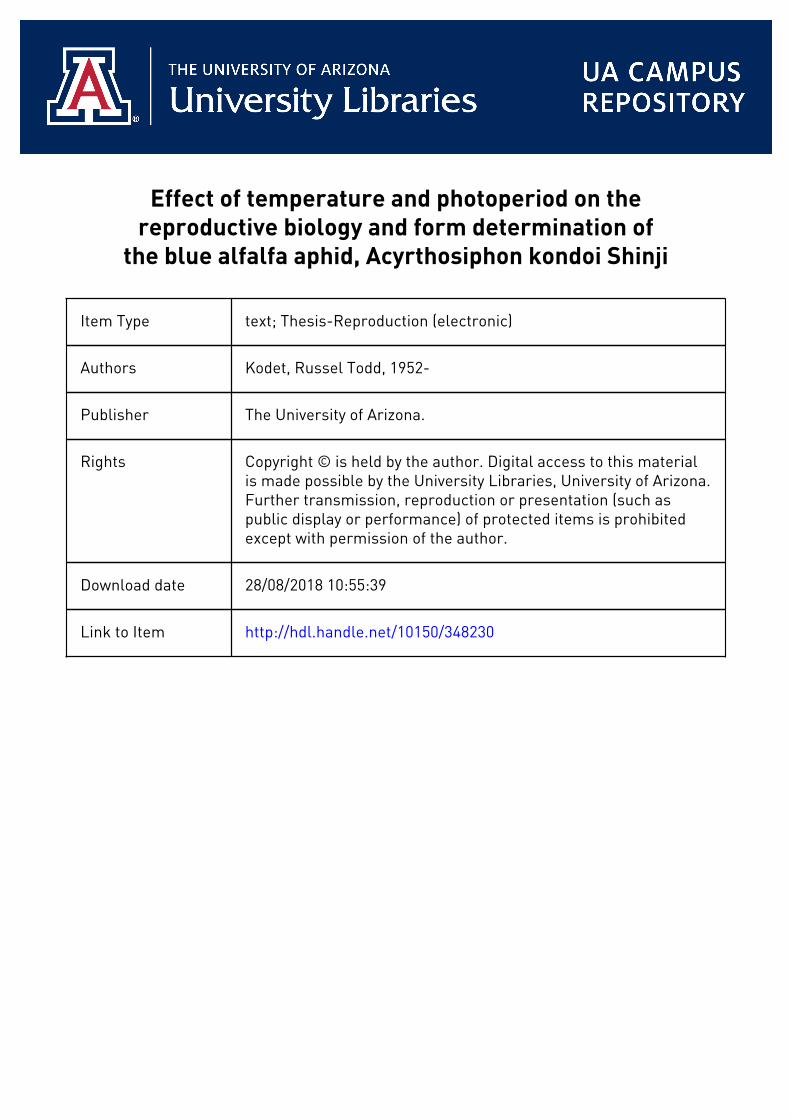
Table 1, The Effect of Temperature and Photoperiod on Nymphal Development of A. kondoi.
Mean Duration (Days)8,Temperature Photophase
(°G) 8 hrgc 12 hrs. l6 hrs.
25 6.31 ab 6.83 b 5*87 a20 7.26 b 8.62 c 6.28 a15 12.60 e 9.68 d 14.00 f10 29.80 i 18.09 g 25.35 h
a Means followed by the same letter are notdifferent at the 0.05 level of significance.

17nymphal period increased with decreasing temperatures at all photophaseso
Treatments which decreased the length of nymphal development decreased the generation time of A* kpndoi and promoted a greater rate of, population increase.
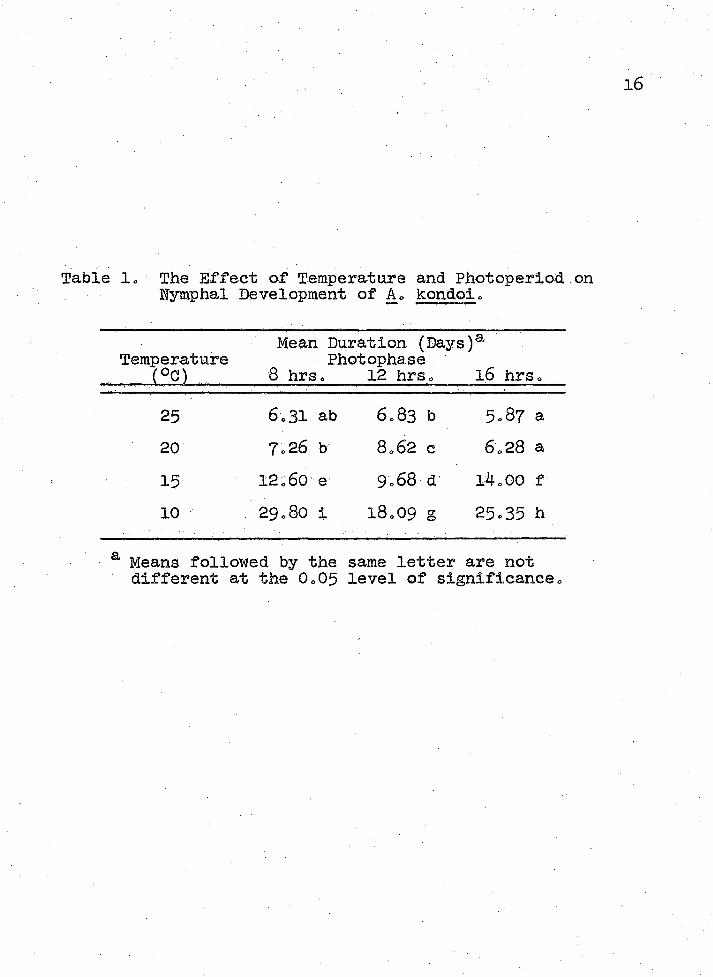
Significantly different accumulations of day-degrees were required for A. kondoi to complete total nymphal development (Table 2)0 An average minimum threshold temperature of,5o40C (found from regression lines for rate of development vs. temperature) was used for day-degree calculations„ These laboratory results indicate the range in which the correct field threshold temperature and day-degree accumulation values are to be found.
The threshold temperature for development of A. kondoi was similar to that of A. pisum (Campbell et al.1974)° Also5 A. kondoi required approximately equivalent numbers of day-degrees to complete development as did A. pisum. However3 since A. kondoi were more numerous than A. pisum early in the spring (Cooperative Plant Pest Report 19769 1977) A. kondoi seem to develop more quickly than A. pisum at lower temperatures. In a pest management decisions the economic threshold for Acyrthosiphon spp. should be lower when A. kondoi predominate since A. kondoi inflicts greater damage to alfalfa than do equal numbers of A. pisum (Nielson and Kodet 1975)° It would be easiest and most
18
Table 2, The Effect of Temperature and Photoperiod on the Mean Number of.Day-Degrees Required for A. kondoi to Complete Nymphal Developmento
Mean Day-Degreesa Temperature Photophase
_____ (°C) 8 hrsc 12 hrs. 16 hrs.
25 135o5 be 133-3 be 115-6 abc20 106.6 ab 125.6 be 92.0 ab15 121.0 be 93-lab 134.4 c10 137-1 c 83-3 a 116.8 be
a Means followed by the same letter are notdifferent at the 0.05 level of significance.

19economical to evaluate both Acyrthosiphon species in alfalfa by defining the economic level in relation to temperature„ The- time consuming process' of Identifying and; separating these two pests could then be avoided by elevating the economic threshold during the ‘warmer months of the growing season when less damaging A. pisum populations predominate„
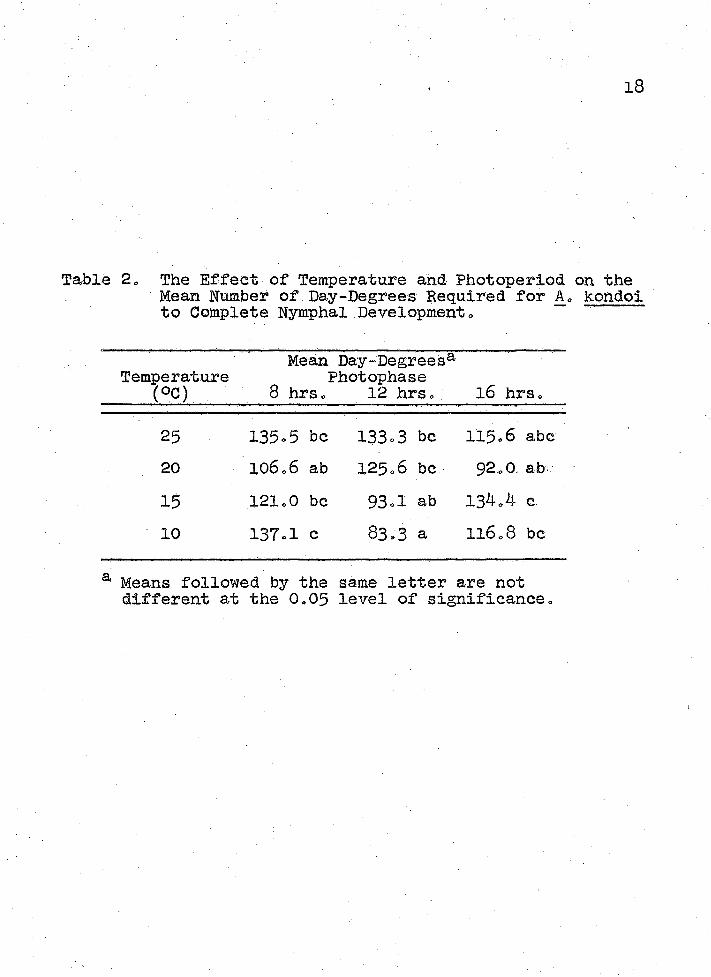
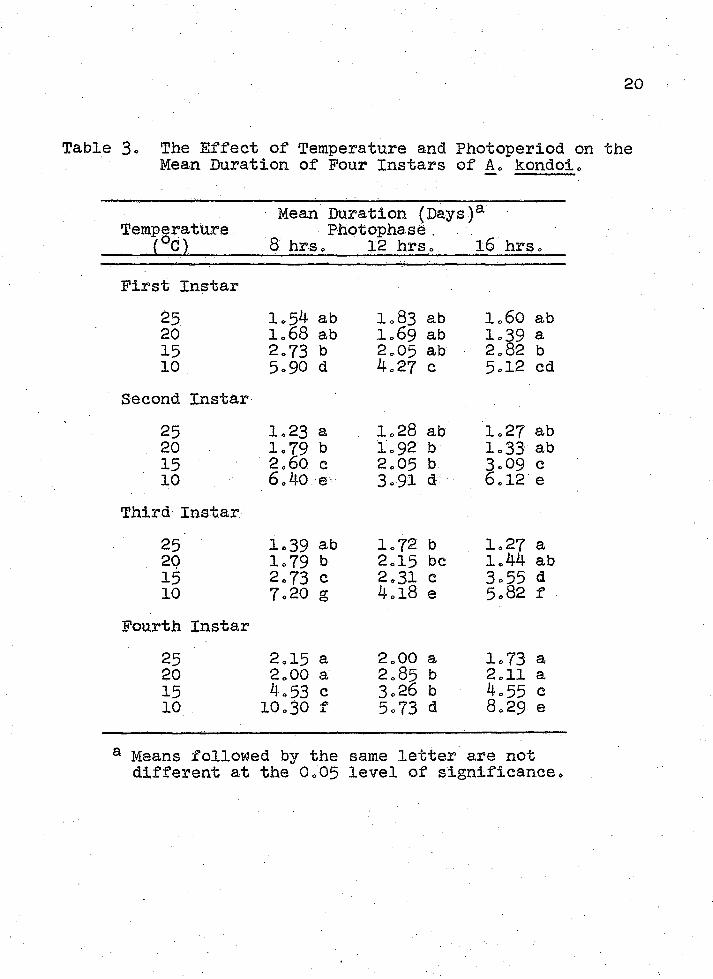
The effect of temperature and photoperiod on the rate of development was not striking for any single instar (Table 3)s but the cumulative temperature/photoperiod interaction effect was strong for the entire nymphal period (Table 1)„The duration of each of the four instars was longer at 10° than at 15°G (Table 3). Slight temperature effects on the duration of instars 2S 3a and 4 were observed between 15° and 20 C5 but measurable differences seldom existed between 20° and 25°C. Furthermore 5 the strong effect of photoperiod observed at 10° on the duration of all four instars was slightly masked at 15°5 was almost hidden at 20° and was completely masked at 25°G <, These results indicated that photoperiod is important in controlling the rate of development of A, kondoi at low temperatures* Photoperiod effects may have acted via the host plant5 Melilotus officionalis 5 or directly upon the aphid. The pathway could not be determined from these experiments =
Progressively more time was required for later instars to complete development^ with the 1st instar
20
Table 3» The Effect of Temperature and Photoperiod on the Mean Duration of Four Instars of A. kondoi„
Mean Duration (Days')8, Temperature Photophase
( C) _____ 8 hrso 12 hrs. l6 hrs«■
First Instar 25 1.5% ab 1.83 ab 1.60 ab20 1.68 ab 1.69 ab 1.39 a15 2.73 b 2.05 ab 2.82 b10 5.90 d 4.27 c 5.12 ed
Second Instar25 1.23 a . 1.28 ab 1.27 ab20 1.79 b 1.92 b 1.33 ab15 2.60 c 2.05 b 3.09 c10 6.4b. ev 3.91 ■CKK- 6.12 e
Third Instar25 1.39 ab 1.72 b 1.27 a20 1.79 b 2.15 be 1.44 ab15 2.73 c 2.31 c 3.55 d10 7.20 g 4.18 e 5.32 f
Fourth Instar 25 2.15 a 2.00 a 1.73 a20 2.00 a 2.85 b 2.11 a15 4.53 c 3.26 b 4.55 c10 10.30 f 5.73 d 8.29 e
a Means followed by the same letter are not different at the 0.05 level of significance.

21
requiring the shortest period of time and the 4th instar the longest period of time (Table 3)» This phenomenon might have been due to the increasingly complex developmental processes that late instars must complete5 i. e,9 ovary maturation, embryo development, or alary muscle formation before becoming adults. The phenomenon might also have been due to the large increase in size between 3rd and 4th in- stars o Nymphs appeared to almost double in size from the 3rd to the 4th instar, while a very small increase occurred between the 1st and 2nd instars»
Nymphal Survivorship Temperature and photoperiod did not interact signif
icantly to affect nymphal survivorship of A. kondoi. The probability of obtaining a greater chi square value than the observed value of 6.74 with 6 degrees of freedom was equal to 0.345.
Analysis of the effect of temperature alone showed that nymphs suffered significantly less mortality at 20°C than they did at any of the other temperatures (Table 4).The observed chi square value of 9.67 with 3 degrees of freedom had a probability of less than or equal to 0.0216. Mortality was approximately equal at 10°, 15° and 25°c.
Significantly greater nymphal survival occurred at 12 hours photophase than at either 8 or 16 hours photpphase (Table 5). The probability of obtaining the observed chi

22
Table 4„ The Effect of Temperature on Nymphal Survivorship of A» kondoi.
Temperature(°C)
Number Surviving Observed Expected ;
Number Dying Observed Expected
25 46,0 47,7 8,0 6.320 50,0 44,5 1.0 6.7
15 45.0 46,9 8,0 6,210 38,0 40,0 8,0 6,1
Table 5» The Effect of Photoperiod on Nymphal Survivorship of A. kondoic
PhotophaseNumberObserved
SurvivingExpected
NumberObserved
DyingExpected
8 hrs. 57.0 61,6 13.0 8,312 hrs. 6l.O 54.3 1,0 8.0l6 hrs. 6i.o 63.1 11,0 8.9

• 23square value of 12085 with 2 degrees of freedom was less than or equal to 0.0016. Moderately high temperatures or intermediate photophases therefore increased survival of Ao kondol nymphs»
Preparturition Period Photoperiod had no effect upon the length of pre-
parturition at 10°, 15° and 20°C (Table 6); but at 25°C preparturition lasted longer at 12 hours photophase than at either 8 or 16 hours photophase.
Temperature significantly affected preparturition by increasing the duration at 15° and 10°G at 8 hours photo- phase, 25°, 150 and 10°c at 12 hours, and at 10oG at 16 hours. Preparturition generally lasted a relatively short period of time when compared with both the parturition and postparturition periods. Acyrthosiphon kondoi appears to have completed extensive ovary development and embryo maturation by the time the final moult occurred in order for parturition to have begun so quickly. The rapid onset of reproduction contributes to the large potential for A. kondoi populations to increase.
Parturition Period Temperatures of 20° and 25°C had no effect upon the
length of the parturition period of A. kondoi at the 3 photo- periods (Table 6). When temperatures were decreased to
24
Table 6 . The Effect of Temperature and Ehotoperiod on theMean Preparturitions Parturition, and Postparturition Periods of A , kondoi«
Mean Duration (Days) Temperature Photdphase
(°C) 8 hrs. 12 hrs.Is - ' .... '
16 hrs.
Preparturition Period25 1.36 a 2.50 be 1.06 a20 0.81 a 0.89 a 1.28 a15 2 0 21 b 1*95 b 1.71 ab10 3.06 c 3*13 c 2.88 c
Parturition Period25 6.21 a 10.33 b 10:59 b20 7*75 ab 8.79 ab 11.53 b15 25.21 c 20.37 c 22.36 c10 . 40.94 d 53*75 e 49.29 e
Postparturition Period25 1.71 a 4.33 a 2.06 a20 0.38 a 1.37 a 2.33 a15 11.79 b 4.21 a 4.93 a10 13.31 b 24.13 c 15*59 b
a Means followed by the same letter are not different at the 0.05 level of significance.

' 2515° and. 10°G5 the parturition period was lengthened considerably .
Photoperiod had no effect on the length of parturition at 15° or 20°C» However, a photophase of 8 hours significantly shortened parturition at 10° and 25°C. This effect, observed at both temperature extremes, might have been caused by adverse growing conditions for the host plant or by disruption of the normal physiology of A. kondoi.
Postparturition Period While the preparturition and parturition periods
were not affected by photophase at 150C, the postparturition period was greatly lengthened by the 8 hour photophase at 15°C and by 12 hours photophase at 10°C. Temperature and photoperiod interactions at 15°/8 hour and 10°/12 hour photo- phases were unclear. An; unknown factor may have exerted less stress on Ao kondoi in these treatments in order for the postparturition period to have increased so dramatically„
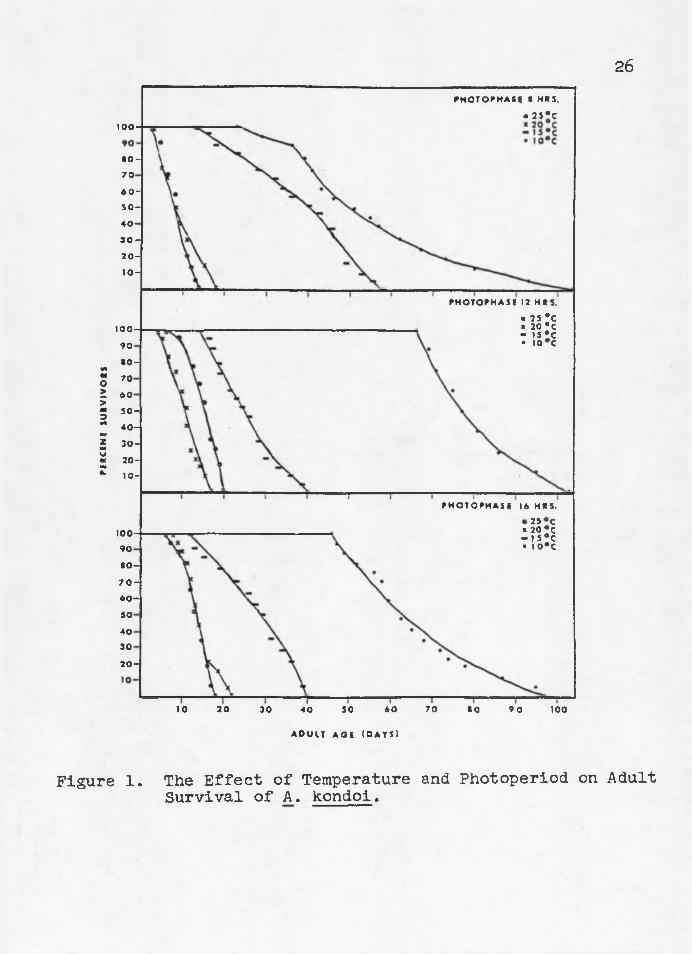
Adult Survivorship Adult survivorship of A» kondoi was approximately
equal at 20° and 25°C but was greatly increased at 10° and 15°C (Figure 1). Fifty percent mortality at 15°C Occurred at day 45 of the adult period at 8 hours photophase, while equivalent mortality at 12 and 16 hour photophases occurred at days 27 and 30, respectively. Fifty percent mortality
Mft
CE
NT
S
UlV
lVO
tS
26FMOTOFMASE • H*S.
• 2 5 *C100
• 0 -7 0
6 0 -
5 0 -
403 0 -
20 -
1 0 -
F M O TO FH AS i 12 H * $ .
• 2 5 *C• 2 0 e C - 15 *C• 1 0 e C
1 0 0
9 0
*0-
706 0
5 0 -
4 0 -
3 0 -
20-
1 0 -
7 H O T O P H A S 6 16 H t S .
25 C
:?i:$• i o e c
100
90• 0-7 0 -
60-
50403 0
20 -
20 7 010 SO 9 03 0 5 0 6 0 1004 0
A O U I T A C E ( D A Y S )
Figure 1. The Effect of Temperature and Photoperiod on Adult Survival of A. kondoi.

at 10°C occurred at day 53 at 8 hours photophasea at day 78 at 12 hours photophase3 and at day 65 at 16 hours photophase„ Despite decreased survivorship at higher temperaturess higher rates of population increase occurred at high temperatures due to increased developmental and fecundity rates 0
FecundityPhotoperiod had no effect on the total fecundity per
female at 15°? 20° or 25°C (Table 7)» However3 A. kondoi produced fewer progeny at long and short photophases at low temperatures (10°C) while A„ pisum produced fewer progeny at high temperatures as photophase increased (Kenten 1955) and produced fewer progeny at intermediate temperatures as photophase decreased (Murdie 1969a., 1969b; Sharmas Larrivee and Theriault 1973S Elliot and McDonald 1976)= This is further evidence that A» kondoi is a "cool weather" aphid since it responded to photoperiods at low temperatures whereas A« pisum responded only to photoperiods at high and intermediate temperatures»
The temperature/photoperiod interaction had no effect on the total fecundity per female of A. kondoi except at 20°/8 hours and 10°/12 hours photophase (Table 7)» Total fecundity was greatly increased by the 10°/12 hour photophase interaction where each female produced an average of 107.6 offspring. Conversely3 total fecundity was reduced at 20°/8 hours.

Table 7= The Effect of Temperature and Photoperiod on the Mean Fecundityper Female and the Mean Fecundity per Female per Day of A, kondol»
Temperature ... (0G)____
Total Fecundity per Female Photophase
8 hrs o 12 hrs, 16 hrs.Daily Fecundity per Female
Photophase 8 hrs. 12 hrs. l6 hrs.
25 26.50 ab* 32.50 ab 38.18 ab 2.60 a 1.93 a 2.93 a20 20.31 a 29.32 ab 46.50 ab 1.94 a 2.64 a 2.72 a15 69.26 b 43.42 ab 47.43 ab 1.73 a 1.64 a 1.6l a10 58.31 ab 107.60 c 69.94 b 1.09 a 1.26 a 0.99 a
'Means followed by the same letter for either total or daily fecundity are not different at the 0„05 level of significance.
oo

29The increased total fecundity per female which oc
curred as temperatures decreased was not highly significant due to large variations in the fecundity between individuals within a treatment. However9 total fecundity per female was in fact increased by lower temperatures (Table 7)® The increase in total fecundity per female at lower temperatures is attributable to the long parturition periods which occurred at lower temperatures (Table S)s since mean daily fecundity of A. kondoi was not significantly affected by the temperature/photoperiod interaction (Table 7). Generally9 longer parturition periods resulted in greater total fecundity.
Photoperiod alone had no effect upon the mean daily fecundity of A, kondoi when it was analyzed separately from temperature. However3 temperature significantly affected daily fecundity. Acyrthosiphon kondoi produced 2.5 offspring daily at 20° and 250CS 1.7 offspring at 15°s and 1.1 at 10°G. The slower daily reproductive rate at lower temperatures did not lower total fecunditys but actually increased total fecundity since A. kondoi reproduced for longer periods of time at these temperatures.
Mean total fecundity of A. kondoi was greatest at 10GC while the greatest total fecundity per female of A. pisum occurred between 20° (Kenten 1955) and 24„4°C (Kilian and Nielson 1971)= Larger populations of A. kondoi than

30A. pisum would, be expected during the cooler parts of the alfalfa, growing, season since A. kondoi develop more quickly than Ao pisum and experience higher fecundity at low temperatures <, These data provide further evidence of the desirability of using a flexible economic threshold based upon temperature when evaluating field populations of Acyrtho- siphon species.
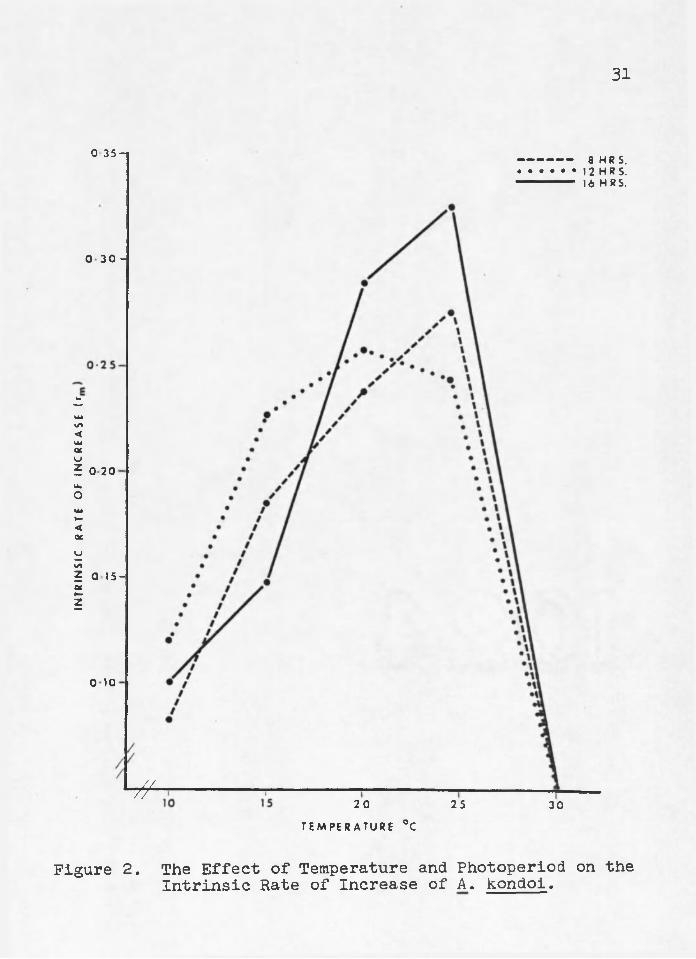
Intrinsic Rate of Increase The intrinsic rate of increase (rm) of A. kondoi was
greatest at 25°/l6 hours photophase and smallest at 10°/8 hours photophase (Figure 2)o The rm increased as temperatures increased from 10° to 20°c at 12 hours photophase and from 10° to 25°C and 8 and 16 hours photophase. The rm fell to zero at all photophases at 30oC where no survival or reproduction occurred after 2 generations.
The rm was greatest at high temperatures despite the fact that total fecundity per female and adult survival were greatest at low temperatures. However3 rm was not based only upon these parameters5 but upon the rate of nymphal development and the daily fecundity as well. The extremely rapid rate of nymphal development at high temperatures overshadowed the success at low temperatures 3 and resulted in a greater rm at higher temperatures.
AcyrthoSiphon kondoi was most numerous early in the growing season when temperatures were cool, and A. pisum was
INT
RIN
SIC
R
AT
E
OF
INC
RE
AS
E
(r31
0 3 5 - 18 H R S .
12 H R S . 16 H R S .
0 3 0 —
0 20
0 1 5 -
0 1 0
//77 20T E M P E R A T U R E ° C
2 5 3 0
Figure 2. The Effect of Temperature and Photoperiod on the Intrinsic Rate of Increase of A. kondoi.

• 32most common later during warmer months (Gopperative Economic Insect Report 1975s Cooperative Plant Pest Report 19765 1977). Therefore? the r^ of A* kpndoi may be greater than that of A., pisum at lower temperatures? although other factors such as disease? predation? and parasitism may differentially affect these two Acyrthosiphon species«
Doubling Time The doubling time of A . kondoi was similar at all
photophases at 20° and 25°G« The doubling time rapidly increased as temperature decreased from 20° to 10°C. The aphid required the most time for the population to double at 10°/8 hours photophase (Table 8).
Form Determination No sexual forms of A. kondpi were produced in these
experiments. Conversely? sexual forms of A. pisum were commonly produced when reared at low temperatures and short photophases (Kenten 1955J Sharma? Larrivee and Theriault 1973; Lamb and Pointing 1975)« Sexual forms are the mechanism by which aphids overwinter in areas with severe win* terso The blue alfalfa aphid may not have been introduced from Eastern Asia but have originated instead from elsewhere in the world where mild winters prevail and the ability to produce sexuals has been lost.

33
Table 8, The Effect of Temperature and Photoperiod on the Doubling Time of A. kondoi.
Mean Doubling Time (Days) Temperature Photophase -
(0C) 8 hrs. 12 hrs. 16 hfs.
25 2 o 52 2.84 2.1320 2.91 2.70 2.3915 3-75 3.06 4.6910 8.62 5.77 7.00

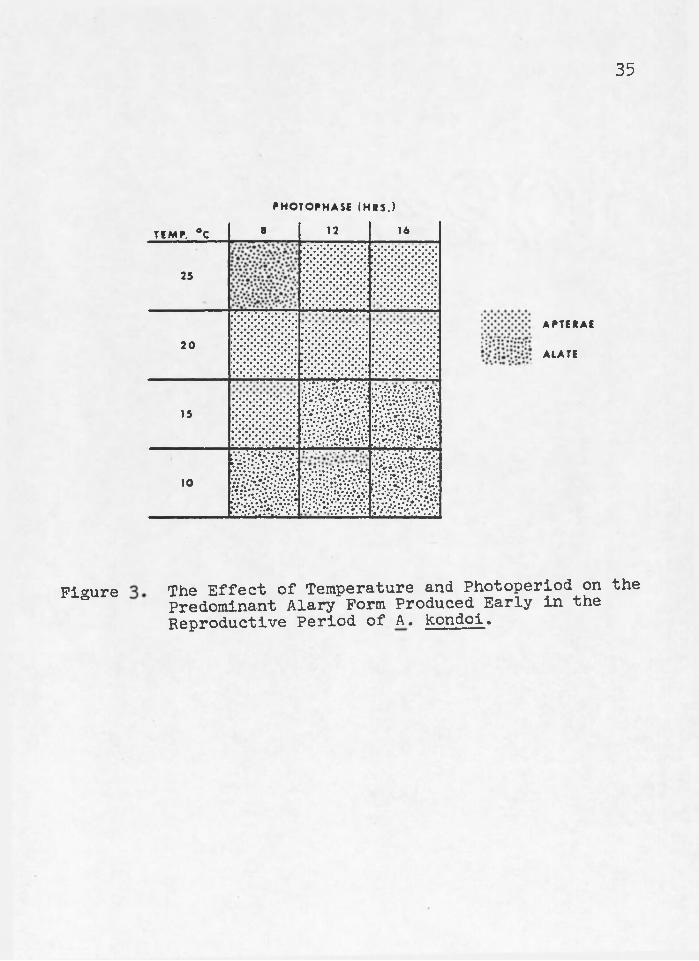
34High temperatures (20 and 25°G) suppressed the pro
duction of alatae5 except at 25°/8 hours where the short photophase produced alataw (Figure 3)« The production of alatae at 25°/8 hours may have been a dispersal mechanism to escape unfavorable environmental, conditions s or a response to an unfavorable nutrient balance in the host plant. High temperatures suppressed alatae production of A. pisum Gregory 1917, Kenten 1955)»
Low temperatures (10° and 15°G) favored the production of alatae5 except at 150/8 hours where apterae were produced in response to the short photophase (Figure 3)° Acyrthosiphon pisum also> produced? apterae;; in response to short photophases at low temperatures (Kenten 1955)«
Acyrthosiphon kondoi switched from the production of alatae or apterae early in the reproductive period (Figure 3) to the production of the opposite respective form later in their reproductive period (Figure 4)s except at 20°/l6 hours where no form reversal occurred. Temperature and photo- period interacted to inhibit form reversal at 20°/l6 hours photophase.
The switch at 25°C occurred near the end of the reproductive period, while at 10°5 15° and 20°C the switch occurred relatively sooner (Table 9)» The switch in the production of alary forms took place after over 70$ of the total reproduction had occurred, except at 10°/12 hours
35
Figure
P H OT O P H A S E ( H R S . )
TEMP. °C 8 12 16
25 111112 0 ■ ■I■115 ille ■10 ■ ■■■
APTERAE
ALATE
The Effect of Temperature and Photoperiod on the Predominant Alary Form Produced Early in the Reproductive Period of A. kondoi.
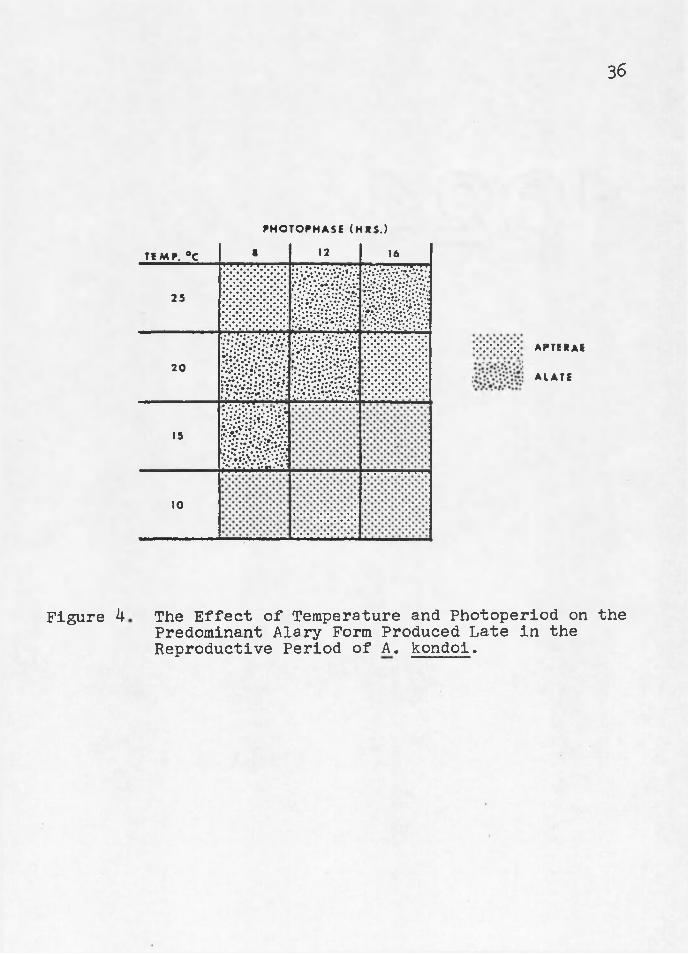
36
Figure 4
P H O T O P H A S E ( H R S . )
T EM P . °C a 12 16
2 5 ■■■2 0 ■*■15
l i
. ....................... .... • •
10# • • • • • • • • •
• • • • • • •
• • • • • • • • • •
• • • • • • l , FeFe1
I : : : : : - : a f t e r a i
ALATE
The Effect of Temperature and Photoperiod on the Predominant Alary Form Produced Late in the Reproductive Period of A. kondoi.
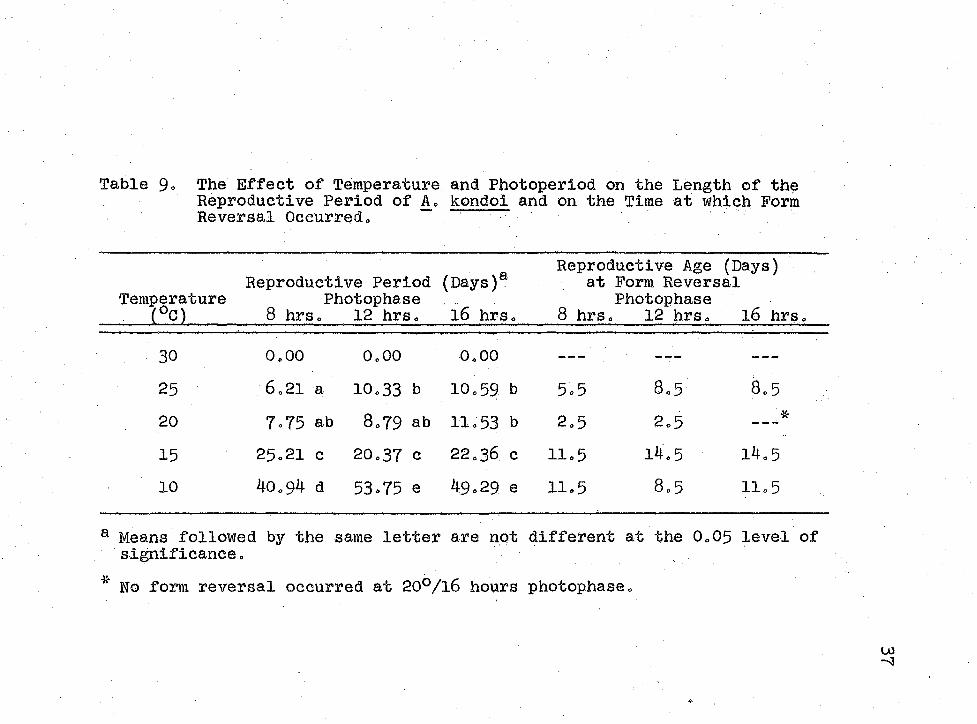
Table 9- The Effect of Temperature and Photoperiod on the Length of the Reproductive Period of A= kondoi and on the Time at which Form Reversal Occurred,
TemperatureReproductive Period (Days)8
Photophase 8 hrs, 12 hrs, 16 hrs.
Reproductive Age (Days) at Form Reversal
Photophase 8 hrs. 12 hrs. 16 hrs.
30 0.00 0.00 0.00 --- — — —
25 6.21 a 10.33 b 10.59 b 5,5 8,5 8.520 7,75 ab 8.79 ab 11.53 b 2.5 2.5 — J*15 25,21 c 20.37 c 22.36 c 11.5 14.5 14.510 40.94 d 53,75 e 49,29 e 11.5 . 8.5 11,5
a Means followed by the same letter are not different at the 0,05 level of significance, .
* Ho form reversal occurred at 20°/l6 hours photophase.

38where 60% of the total reproduction was completed (Table 10). Little difference existed between the times required for A. kondoi to produce 70$ of their total offspring at 20° and 25°C. Generallys this period of time was considerably lengthened by Idwering temperatures to 15° and 10°C. Therefore^ a majority of reproduction occurred in a relatively short period of time and before the switch had occurred. Further5 a treatment may be classified as alatae- or apterae-promoting by observing the predominant alary form produced early in the reproductive period. Those forms produced after the switch had occurred constituted only a small percentage.of the alary forms produced in a treatment, Kenten (1955) showed that A. pisum switched from the production of females early in the reproduetive period to the production of males later in the reproductive period,, but she did not observe similar changes in the alary forms of viviparous A. pisum.
The significance of the switching phenomenon may be that it is an inherent genetic mechanism by which A. kondoi insure that both alary forms are present at all times in the population to guard against sudden changes in environmental conditions which could eliminate all of one form. The mechanism may also insure that alates are always present to disperse and colonize new food sources.

39
Table 10. Reproductive Age at which 70$ of the Total Reproduction of A. kondoi Had Occurred.
Temperature(°c) ..
Photophase (Hours) 8 hrs. 12 hrs. 16 hrs.
30 . — — — — — —
25 3.3 3.8 3.520 ■ 3.5 3.0 3.515 5.3 3.7 5.810 11.0 9.8 9.0

CONCLUSIONS
The upper threshold temperature for development of A, kondoi ranged between 25° and 30°C since supernumary moults occurred at 25°/12 hours and no survival occurred at 30°C. Decreasing temperatures from 25° to 10°C increased the length of nymphal development. An intermediate photo- phase (12 hours) increased the developmental period at high temperatures3 but decreased the developmental period at low temperatures^ The temperature/photoperiod interaction on nymphal development was strongest at low temperatures (10°C) while high temperatures (20° and 25^C) masked the effect of photoperiod on the nymphal stage. Early instars required less time to complete development than did late instars. Nymphal survivorship was optimum at 20oC and at 12 hour photophases.
Temperature and photoperiod interacted at low temperatures to influence preparturition, parturition, and post- parturition periods, while high temperatures masked the effects of photoperiod. Lower temperatures increased the lengths of preparturition, parturition, and postparturition periods. Total fecundity per female increased with decreasing temperatures; conversely, daily fecundity per female increased with increasing temperatures. The optimum
40

temperature for total fecundity of A. kondoi (10°C) was lower than that of A» pisum (20°C)« Temperature and photo- period interacted only very slightly to affect fecundity of A. kondoi* whereas these interactions strongly affected the fecundity of A. pisum.
The intrinsic rate of increase rose with increasing temperature9 while the doubling time decreased with increasing temperatures. The optimum conditions for A. kondoi populations to increase mas 25°/l6 hours photophase = Acyrtho- siphon kondoi was generally better adapted to cooler growing conditions than A, pisum. A flexible economic threshold based on temperaturesis: needed: to evaluate these pests of alfalfa.
Sexual forms of A. kondoi were not produced as the result of simple temperature/photoperiod interactions. The conditions that will initiate the development of sexuals of this species are unknown. High temperatures promoted the development of apterous forms of A. kondoi9 while low temperatures produced predominantly alatae. High temperatures also suppressed the development of alatae of A. pisum. Acyrthosjphon kondoi switched from the production of alatae or apterae early in their reproductive period to the production of the opposite alary form5 respectively, later in their reproductive period. The mechanism of this phenomenon is unknown.

SUMMARY
The blue alfalfa aphid was introduced into California in 19?4 and caused severe damage to alfalfa in several Western States„ The biology of the aphid was unknown. Therefore5 studies were done to determine the effects of temperature and photoperiod on the biology and form determination of Acyrthosiphon kondoi,
Aphids were caged individually on Melilotus. officion- alis (L.) Lam, and reared in environmental chambers at 5 constant; temperatures.. and 3 photophase s . Developmental, fecundity, and survival: data were recorded daily.
Developmental rates increased as temperatures rose from 10° to 25°C, while no survival occurred at 30oC, Late instars required progressively more time to complete development than did early instars. The developmental rate was greatest at 25°CS while survival was optimum at 20oC. A 12 hour photophase was optimum for both nymphal development and survival. High temperature^ masked the effect of photo- period on nymphal development,
Decreasing temperatures increased the length of pre- parturition, parturition, and postparturition periods. Moderate and high temperatures masked the effect of photoperiod on all of the above adult parameters. Photoperiod effects
42

43were observed at low temperatures. Total fecundity was greatest at low temperatures while dally fecundity was greatest at high temperatures,
The optimum conditions for increase of A. kondoi populations was at 250/l6 hours„ The ihtrihsic rate of increase decreased with decreasing temperatures.
Ho sexual forms of A. kondoi were produced. High temperatures promoted the development of apterous forms5 while low temperatures favored the development of alatae forms. Adults switched from the production of one alary form early in their reproductive period to the production of the opposite alary form later in their reproductive period. Photoperiod affected form determination at 25° and 150C.

LITERATURE CITED
Ackerman5 L. 1926. The Physiological Basis of Wing Production in the Grain Aphiti-,, J„ Exp, Zool. 44:l-6l.
Birch, L, C, 19480 The Intrinsic Rate of Natural Increase of an Insect Population. J. Animal Ecol. 17;15-26.
Call, L. E. 1918. Climate and Injurious Insect Investigations. Kansas Agric. Exp. Sta. Rpt. 1917-1918:20-21.
Campbell, A., B. B. Frazer, N. Gilbert, A. P. Gutierrez, and M. MacKauer. 1974. Temperature Requirements of Some Aphids, and Their Parasites. J. Appl. Ecol. 11:431- 438.
Cooperative Economic Insect Report, 19750 S. D. A. Animaland Plant Health Inspection Service Vol. 25.
Cooperative Plant Pest Report. 1976. U. S. D. A, Animal and Plant Health Inspection Service Vol. 1.
Cooperative Plant Pest Report. 1977. Uo s. D. A. Animal and Plant Health inspection Service Vol. 2.
Dickson, A. F. G. and D. M. Glenn. 1971. Morph Determination in the Bird Cherry-oat Aphid, Rhopalosiphum padi L. Ann. Appl. Biol. 68:11-21.
Dickson, R. C. 1975. Identity, Origin and Host Range of the Blue Alfalfa Aphid. Proc. Fifth Calif. Alfalfa Symp.pp. 22-23.
Eastop, V. F. 1971. Keys for the Identification of Acyrtho- siphon (Hemiptera: Aphidldae). Bull. Brit. Museum (Nat. Hist.) Entomol. 26:3-115.
Elliot, H. J. and F. J. D. McDonald. 1976. Reproduction of a Parthenogenetic Aphid Aphis craccivora Koch: Embryology, Ovarian Development and Fecundity of Apterae and Alatae. Austral. J. Zool. 24:49-63.
Ewing, H. E, 1916. Eighty-seven Generations in a Partheno- . genetic Line of Aphis avenae Fab. Biol. Bull.,Woods Hole 31s53-112.
44

45Gaylors M. J«3 and W, L« Sterling. '1975« Effects of Temper
ature on the Developments Egg Production, and Survival of the Cotton Fleahopper. Pseudatomoscelis serlatus. Environ. Entomol. 4:487-490.
Gregory, L. H. 1917° The Effect of Starvation on WingDevelopment of Macrosiphum destructor. Biol. Bull., Woods; Hole 33; 296-303°
Hille His Lumbers, D. 1966. Polymorphism in Aphididae. Ann. Rev. Entomol. 11;47-78.
Johnson, S. 1966. Wing Polymorphism in Aphids. IV. TheEffect of Temperature and Photoperiod. Ent. Exp. and Appl. 9;301-313°
Kenten, J. 1955° The Effect of Photoperiod and Temperature On Reproduction in Acyrthosiphon pisum (Harris) and on the Forms Produced. Bull. Entomol. Res. 46; 599-624.
Kilian> L° and M. W. Nielson. 1971° Differential Effects of Temperature on the Biological Activity of Four Bio- types of the Pea Aphid. J. Econ. Entomol. 64;153-155°
Kitzmiller, J. B. 1950. The Time Interval between Determination and Differentiation of Wings, Ocelli, and Wing Muscles in the Aphid Macrosiphum sanborni (Gillette). Am. Nat. 84;23-50.
Kono, T. 1975° Distribution and Identification of the Blue , Alfalfa Aphid. Proc. Fifth Calif. Alfalfa Symp. pp. 24-27.
Lamb, K. P. and D. White. 1966. Effect of Temperature, Starvation and Crowding on Production of Alate YOung by the Cabbage Aphid, Brevjcoryne brassicae. Ent. Exp. and Appl. 9:179-184.
Lamb, R. J. and P. J. Pointing. 1975° The ReproductiveSequence and Sex Determination in the Aphid, Acyrthosiphon pisum. J. Insect Physiol. 21;1443-1446.
Lees, A. D. 1959° The Role of Photoperiod and Temperature in the Determination of Parthenogenic and Sexual Forms in the Aphid Megoura viciae Buckton. I. The Influence of these Factors on Apterous Virginoparae and their Progeny. J. Insect Physiol. 3:92-117°

46Lees5 Ao D. 1963» The Role of Photoperiod and Temperature in the Determination of Parthenogenic and Sexual Forms in the Aphid Megoura vieiae Buckton. III. Further Properties of the Maternal Switching Mechanism. Jo Insect Physiolo 9sl53-l64»
Lees 5 A o Do 1964» The Location of the PhotoperiOdic Receptors in the Aphid Megoura viciae Buckton. J« Exp. Biol. 41:119-133, " "
Lees^ A. D, 1966. The Control of Polymorphism in Aphids.Adv. Insect Physiol. 3:207-277.
Leess A. Do 1967= The Production of the Apterous and Alate Forms in the Aphid Megoura vieiae Buckton3 with Special Reference to the Role of Crowding. J. Insect Physiol. 13;289-3180
MacG-illivarys M. E. and G. B. Anderson. 1964. The Effect of Photoperiod and Temperature on the Production of Gamic Forms in Macrosiphum euphorbias (Thomas).Can. J. Zool. 42:491-510.
Marcovitch, S. 1924. Migration of the Aphidiae and the Appearance of Sexual Forms as Affected by the Relative Length of Daily Light Exposure. J. Agric. Res. 27:513-522.
Messenger3 P. S. 1964. The Influence of Rhythmically Fluctuating Temperatures on the Development and Reproduction of the Spotted Alfalfa Aphid5 Therjoaphis maculata. J. Econ. Entomol. 57;71-76. .
Miyassakig M. 1971« A Revision of the Tribe Macrosiphini of Japan. Insecta Matsum. 34:1-247=
Murdies G, 1969a. The Biological Consequences of Decreased Size Caused by Crowding or Rearing Temperatures on Apterae of the Pea Aphid9 Acyrthosiphon pisum (Harris). Trans. R. Ent. Soc. Lond. l2l:443-455.
Murdie3 G. 1969b. Some causes of size Variation in the Pea Aphids Acyrthosiphon pisum. Trans. R. Ent. Soc. Lond. 121:423-442.
Nielsons M. W. and R. T. Kodet. 1975» The Blue AlfalfaAphid in Arizona.Proc. Fifth Calif. Alfalfa Symp. pp. 39-40.

4?Nielson3 M« W .5 L. Moore3 arid R. T. Kodet. 1976 = Far Eastern
Pest Invades Arizona Alfalfa Fields« Prog. Agric. in Az. 28:18-19.
Reinriard, H. J. 1927« The Influence of Parentage3 Nutritions.Temperature and Crowding on Wing Production in Aphis gossypii Glover. Bull. Tex. Agric. Exp. Sta. 353: 1-19,
Schaeferss G. A. and F. D. Judge. 1971« Effects of Temperature s Photoperiod3 and Host Plant on Alary Polymorphism in the Aphid3 Chaetosiphon fragaefolil.J. Insect Physiol. 17:365-379°
Sharmas M. L. 3 J. M. Larrivee and L. M. Theriault. 1973°Effets de la Photoperiode et des Temperatures Moy- ennes de 15°C sur la Fecondite et la Production des Sexues Chez le Puceron du Pois3 Acyrthosiphon pisum (Aphididae: Homoptera). Can. Ent.105:947-956.
Sharmas R . .K ° 3 V. M. Stern and R. W. Hagemann. 1975° Blue Alfalfa Aphid Damage and its Chemical Control in the Imperial Valley. Proc. Fifth Calif. Alfalfa Symp. pp. 29-30.
Shinjis J. 0. and T. Kondo. 1938. Aphididae of Manchoukuo with the Description of Two New Species. Kontyu 12: 55-69°
Shulls A. F. 1942. The Mechanism through which Light andHeat Influence Genetic Factors for Wing Development 1 in Aphids. J. Exp. Zool. 89:183-195°
Siddiquij W. H.s C._A. Barlow and P. A. Randolph. 1973°Effects of Some. Constant and Alternating Temperatures on Population Growth of the Pea Aphid9 Acyrthosiphon pisum (Homoptera: Aphididae). Can. Ent. 105: I4F I 56:----
Smiths L. M. 1937° Growths Reproductions Feeding and Wing , Development. of the Mealy Plum Aphid in Relation to Climatic Factors. J. Agric. Res. 54:345-363°
Snedecors G. W. and W. G. Cochran. 1967° Statistical Methods9 6th ed. Arne S 3 . Iowa State University Press. 593 PP°
Takahashis R. 1965° Some New and Little Known Aphididaefrom Japan (Homoptera). Insecta Matsum. 28:19-61.

48Tao, G.
Wadley,
White,
Wilson,
C. 1963. Revision of Chinese Macrosiphinae. Bull. Plant Prot., Taiwan 5:162-205.F. M. 1923. Factors Affecting the Proportion of Alate and Apterous Forms of Aphids. Ann. Ent. Soc. Am. 16:279-303.
W. S. 1946. The Environmental Conditions Affecting the Genetic Mechanism of Wing Production in the Chrysanthemum Aphid. Am. Nat. 80:245-270.F. 1938. Some Experiments on the Forms of Aphis chloris Koch (Aphididae). Trans. R. Ent. Soc. Bond. 67:165-180.

m


















