
, Colin Meyer , Keith W. Ward , Mark E. Cooper , Judy B ...
44
1 | Page A derivative of Bardoxolone methyl, dh404, in an inverse dose-dependent manner, lessens diabetes-associated atherosclerosis and improves diabetic kidney disease. Sih Min Tan 1 , Arpeeta Sharma 1 , Nada Stefanovic 1 , Derek Y.C. Yuen 1 , Tom C. Karagiannis 2 , Colin Meyer 3 , Keith W. Ward 3 , Mark E. Cooper 1 , Judy B. de Haan 1 1 Oxidative Stress Laboratory, Diabetic Complications Division, Baker IDI Heart and Diabetes Institute, Melbourne, Australia 2 Epigenomic Medicine, Baker IDI Heart and Diabetes Institute, Melbourne, Australia 3 Reata Pharmaceuticals Inc. 2801 Gateway Dr, Irving, TX 75063, United States. Running title: dh404 improves end-organ injury in diabetic mice Corresponding author: Dr Judy B. de Haan Address: 75 Commercial Road, Melbourne, Victoria 3004, Australia Fax number: +61(3) 8532 1100 Phone number: +61(3) 8532 1520 Email address: [email protected] Word count of abstract: 200 Word count of main text: 3996 Number of figures: 6 Number of tables: 2 References: 44 Page 1 of 44 For Peer Review Only Diabetes Diabetes Publish Ahead of Print, published online April 16, 2014
Transcript of , Colin Meyer , Keith W. Ward , Mark E. Cooper , Judy B ...

1 | P a g e
A derivative of Bardoxolone methyl, dh404, in an inverse dose-dependent manner,
lessens diabetes-associated atherosclerosis and improves diabetic kidney disease.
Sih Min Tan1, Arpeeta Sharma
1, Nada Stefanovic
1, Derek Y.C. Yuen
1, Tom C.
Karagiannis2, Colin Meyer
3, Keith W. Ward
3, Mark E. Cooper
1, Judy B. de Haan
1
1Oxidative Stress Laboratory, Diabetic Complications Division, Baker IDI Heart and
Diabetes Institute, Melbourne, Australia
2 Epigenomic Medicine, Baker IDI Heart and Diabetes Institute, Melbourne, Australia
3Reata Pharmaceuticals Inc. 2801 Gateway Dr, Irving, TX 75063, United States.
Running title: dh404 improves end-organ injury in diabetic mice
Corresponding author:
Dr Judy B. de Haan
Address: 75 Commercial Road, Melbourne, Victoria 3004, Australia
Fax number: +61(3) 8532 1100
Phone number: +61(3) 8532 1520
Email address: [email protected]
Word count of abstract: 200
Word count of main text: 3996
Number of figures: 6
Number of tables: 2
References: 44
Page 1 of 44
For Peer Review Only
Diabetes
Diabetes Publish Ahead of Print, published online April 16, 2014

2 | P a g e
Abstract
Oxidative stress and inflammation are inextricably linked and play essential roles in
the initiation and progression of diabetic complications such as diabetes-associated
atherosclerosis and nephropathy. Bolstering antioxidant defences is an important
mechanism to lessen oxidative stress and inflammation. In this study, we have used a
novel analogue of the Nrf2 agonist bardoxolone methyl, dh404, to investigate its
effects on diabetic macrovascular and renal injury in STZ-induced diabetic ApoE-/-
mice. We show that dh404, at lower but not higher doses, significantly lessens
diabetes-associated atherosclerosis with reductions in oxidative stress (in plasma,
urine and vascular tissue) and pro-inflammatory mediators TNF-α, ICAM-1, VCAM-
1 and MCP-1. We demonstrate that dh404 attenuates functional (urinary
albumin/creatinine ratio) and structural (mesangial expansion) glomerular injury, and
improves renal tubular injury. Liver functional and structural studies showed that
dh404 is well tolerated. Complementary in vitro studies in NRK cells showed that
dh404 significantly upregulates Nrf2-responsive genes, HO-1, NQO1 and GSH-S
transferase, with inhibition of TGFβ-mediated pro-fibrotic fibronectin, collagen I and
pro-inflammatory IL-6. Higher doses of dh404 were associated with increased
expression of pro-inflammatory mediators, MCP-1 and NF-κB. These findings
suggest that this class of compound is worthy of further study to lessen diabetic
complications but that dosage needs consideration.
Page 2 of 44
For Peer Review Only
Diabetes

3 | P a g e
Introduction
Diabetes mellitus is a common risk factor for both chronic kidney disease and
atherosclerosis[1,2]. Despite the use of conventional therapies that include blood
pressure, glucose lowering and hyperlipidemic treatments, significant numbers of
diabetic patients suffer morbidity or mortality as a consequence of cardiovascular
complications and/or progress to end stage renal failure. Novel interventions are
needed to satisfy this unmet clinical need to limit multiple diabetes-associated
complications.
Evidence strongly supports a role for oxidative stress and inflammation as
underlying pathogenic mechanisms of both diabetic renal and atherogenic
complications. With oxidative stress and inflammation now recognised to be
inextricably linked[3], approaches that limit oxidative stress are likely to translate to
reduced inflammation. Limiting oxidative stress by bolstering antioxidant defences is
a novel alternative approach to antioxidant therapy that may yield more efficacious
outcomes than standard vitamin therapy[4]. One approach that has attracted
significant attention is the augmentation of antioxidant defences via an upregulation
of the NFE2-related factor 2 (Nrf2)/Keap-1 pathway[5-9].
Numerous small molecule activators of the Nrf2/Keap1 pathway have been
identified [10,11] including bardoxolone methyl (BM), also known as CDDO-methyl,
a potent synthetic triterpenoid inducer of the phase 2 response[12,13]. Bardoxolone
methyl and other related antioxidant inflammation modulators (AIM) such as CDDO-
imidazole (RTA 403) have demonstrated structural and functional improvements in
several rodent models of renal disease[14-16], along with an increase in protective
Nrf2, PPARγ and HO-1 genes[16]. Given the known negative effects of oxidants and
inflammation on the vasculature, and the promising antioxidant, anti-inflammatory
and tissue-protective effects of AIM compounds[13], we hypothesised that Nrf2
agonists may additionally protect the diabetic vasculature against accelerated
atherosclerosis. Our hypothesis is strengthened by recent data showing an adaptive
induction of Nrf2-driven antioxidant genes in response to hyperglycaemia in human
endothelial cells[17] and the use of the Nrf2 activator, sulforaphane, to induce Nrf2
activation of downstream target genes in endothelial cells[18].
Interest in BM was further enhanced by two recent Phase 2 clinical trials
where this compound showed a dose-dependent increase in the estimated glomerular
Page 3 of 44
For Peer Review Only
Diabetes

4 | P a g e
filtration rate (eGFR), initially after 56 days of treatment[19] and in a second study
(BEAM) after 24 weeks of treatment in patients with advanced chronic kidney disease
and type 2 diabetes[20]. In the latter study, the positive effects of drug treatment
persisted at 52 weeks after removal of the drug at 24 weeks. However, issues of drug
toxicity and off-target effects of this drug-class have been raised, particularly after the
larger Phase 3 clinical trial (BEACON) with primary endpoints of time-to-first
occurrence of either end-stage renal disease or cardiovascular death[21] was
terminated prematurely due to unexplained adverse cardiovascular events. Recently
the results of BEACON have been published and suggest a potential effect of BM on
heart failure further emphasising that this drug class needs more extensive
investigations with respect to not only renal but also cardiovascular endpoints[22].
Potential problems with toxicity and a worsening of diabetic nephropathy were also
raised in a preclinical study examining the effects of BM analogues, RTA 405 and
dh404, on renal outcomes in diabetic Zucker fat rats[23]. The issue of toxicity was
subsequently addressed in a follow-up study which showed that RTA 405 and dh404
are well tolerated and did not confirm the previous report of hepatic and renal
toxicity[24].
To further clarify the effects of Nrf2 activation on diabetic renal injury and to
investigate its potential role in lessening diabetes-associated atherosclerosis, in this
study we have treated diabetic ApoE-/- mice with the BM analogue, dh404, for 18
weeks and have assessed parameters associated with renal structural and functional
injury, as well as atherosclerosis. As rodents metabolize BM to a metabolite which
itself is not toxic to higher mammals but is quite toxic to rodents, BM itself cannot be
used for chronic rodent studies. Therefore tool compounds, including dh404 (also
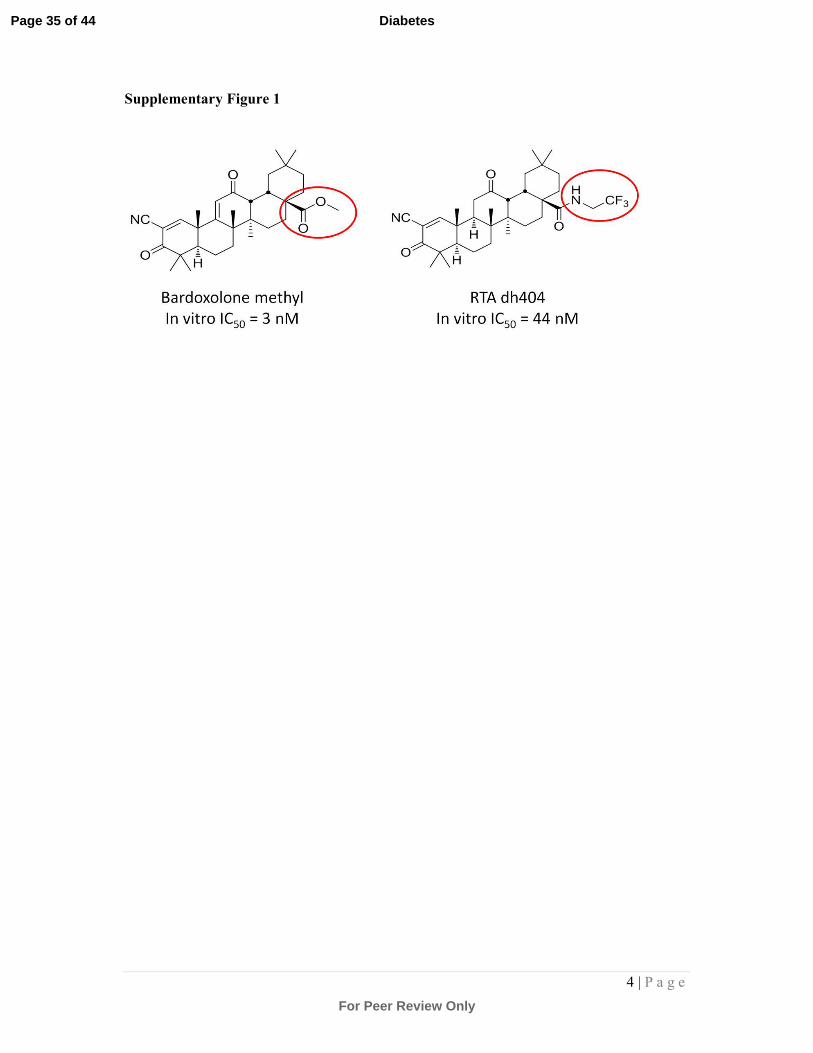
known as dh-CDDO-trifluoroethyl amide, see supplementary Fig.1), are used to probe
rodent pharmacology for this class and inform on the likely clinical behaviour of BM.
Indeed, BM and dh404 exhibit very similar qualitative biological properties [25] and
the same mechanism of action, where dh404 has been shown to activate Nrf2 by
preventing its degradation as a result of interrupting the ability of Keap1 to bind to
Nrf2. Therefore, dh404 is an appropriate surrogate for studying this class of
compound. We now present evidence for the first time, that dh404 lowers oxidative
stress, pro-inflammatory mediators and protects against diabetes-associated
Page 4 of 44
For Peer Review Only
Diabetes

5 | P a g e
atherosclerosis. In addition, dh404 improves renal structural and functional injury
while showing no detrimental effect on liver function in this diabetic model.
Materials and Methods
Bardoxolone methyl derivative, dh404
Dh404, solubilized in sesame oil (Spectrum Chemical MFG. Corp.) at 3, 10 or
20mg/kg per body weight, was administered to mice once per day by oral gavage.
Animals
Eight-week old male C57Bl/J6 ApoE-/- mice purchased from the Animal Resource
Centre (Perth, Australia) were rendered diabetic by two intraperitoneal injections of
streptozotocin (STZ, Sigma, USA) at 100mg/kg/day on two consecutive days[26-28].
In a first cohort, 10-week old diabetic mice were gavaged with either sesame
oil (SO) or 10 or 20mg/kg dh404. Additionally, a group of non-diabetic mice received
20mg/kg dh404. Aortas and kidneys were collected for qRT-PCR after 5 weeks of
gavage. A second cohort of diabetic mice were gavaged with SO or 3, 10 or 20mg/kg
dh404 for 18 weeks. Additionally, sham-injected non-diabetic mice were gavaged
with 20mg/kg dh404. Weekly blood-glucose, body-weights and blood-pressure
measurements (using non-invasive tail-cuff plethysmography[29]) were performed.
Three days prior to termination, urine was collected in 24h metabolic cages. Mice
were killed by lethal injection of 2,2,2-tribromoethanol (Sigma, St Louis, MO, USA)
and direct puncture of the right ventricle to obtain blood. Plasma was stored at -80oC.
Plasma and urine were analysed for plasma ALT, AST and urine creatinine
(Australian Specialised Animal Pathology Laboratory, Mulgrave, Victoria, Australia).
Aortas, kidney and livers were fixed in 10% neutral buffered formalin. Kidney cortex
was snap frozen in liquid nitrogen for RNA extraction.
Evaluation of Atherosclerotic Lesion
En face analysis of lesions was conducted after staining with Sudan IV-Herxheimer’s
solution[26-28]. Plaque area was calculated as the proportion of aortic intimal surface
area occupied by red stained plaque (Adobe Photoshop v6.0.1; Adobe Systems, NSW,
Australia).
Page 5 of 44
For Peer Review Only
Diabetes

6 | P a g e
Lesions within the sinus were visualised after staining with Oil Red O and
quantitated as described previously[26-28].
Kidney Function Assessment
Urinary albumin to creatinine ratio was measured using a mouse albumin ELISA kit
(Bethyl Laboratories, Montgomery, TX, USA). Plasma cystatin C was measured
using a mouse cystatin C ELISA kit[30](BioVendor, Brno, Czech Republic).
Oxidative Stress Assessment
Urinary 8-isoprostane and 8-hydroxy-2-deoxyguanosine (8-OHdG) were assessed
using commercially available EIA kits (Cayman Chemical, USA and StressMarq
Biosciences Inc, Canada, respectively). Measures were expressed relative to urinary
creatinine. Plasma was analysed for diamicron reactive oxygen metabolites (dROMs)
particularly hydroperoxides[31] using a FRAS-4 system and expressed in Carratelli
Units[32].
Histological assessment of kidney
Mesangial and tubulointerstitial area (TIA). Kidney sections were stained with
periodic-acid Schiff (PAS). Mesangial area was quantitated using Image-Pro Plus
v6.0 (Media Cybernetics, USA) and expressed as percentage of PAS-stained area per
glomerular cross sectional (gcs) area. TIA was assessed using a point-counting
technique[33].
Glomerulosclerosis. The degree of sclerosis in each glomerulus was graded on a scale
of 0-4[34].
Histological assessment of liver
Livers were stained with haematoxylin and eosin, and scored blinded on a scale from
0-5 to determine hepatocellular cytoplasmic rarification and periportal
inflammation[19]. Five to eight livers were examined per group. Ten sections were
scored and averaged to obtain an overall score for each liver.
Page 6 of 44
For Peer Review Only
Diabetes

7 | P a g e
Immunohistochemistry
Four-µm aorta paraffin sections were stained for nitrotyrosine, 4-hydroxynonenal (4-
HNE) and VCAM-1. Four-µm kidney sections were stained for collagen IV and
nitrotyrosine[27,28].
Sections were examined under light microscopy (Olympus BX-50; Olympus
Optical) and digitised with a high-resolution camera. Digital quantifications (Image-
Pro Plus,v6.0) were performed in a blinded manner.
Quantitative Reverse Transcription-Polymerase Chain Reaction (qRT-PCR)
RNA from kidney and aorta was extracted using TRIzolR Reagent and cDNA
synthesised as described previously[27,28]. Gene expression was determined after
real-time quantitative RT-PCR and analysed as described previously[27,28]. Gene
expression was normalised relative to 18S ribosomal RNA. Primers and probes are
shown in Supplementary Table 1.
In vitro experiments in normal rat kidney (NRK) cells
Tubular NRK52E cells (ATCC, Rockville, USA) were maintained in Dulbecco's
modified Eagle medium containing 10% serum and 25mmol/l glucose. Cells were
serum starved and pre-treated with dh404 at 0.25µM, 0.5µM, 0.75µM or vehicle
(DMSO) for 4h before stimulation with 10ng/ml TGF-β1 for 72h[35]. Treatment at
1µM dh404 and above for 48hr resulted in significant cell death (Supplementary
Figure 2). Cells were collected for qRT-PCR or Western blot analysis[28].
Statistical Analyses
Data are expressed as mean±standard error of mean (SEM) and analysed by one-way
ANOVA with Newman-Keuls post hoc testing. Statistical analyses were performed
using GraphPad Prism version6.0 (GraphPad Software, La Jolla, CA, USA). P<0.05
was considered statistically significant.
Results
Metabolic Parameters
Dh404 had no effect on systolic blood pressure in diabetic mice (diabetic mice: 129±2
vs 129±1; 130±1; 134±1 mmHg for 3, 10, 20mg/kg dh404 respectively). Body
Page 7 of 44
For Peer Review Only
Diabetes

8 | P a g e
weights of STZ-diabetic mice were significantly lower than non-diabetic controls
(Table 1). However, diabetic mice treated with lower doses of dh404 (3 and
10mg/kg/day) exhibited higher body-weights compared with both vehicle-treated
diabetic mice and diabetic mice treated with the highest dose of 20mg/kg dh404.
Vehicle-treated diabetic mice also displayed significantly higher kidney, liver and
lung to body-weight ratios compared with non-diabetic mice. Treatment of mice with
3 and 10mg/kg/day dh404 attenuated the increase in kidney to body-weight ratio
associated with diabetes (Table 1). Heart to body-weight ratios were unaffected by
diabetes and dh404 treatment. All diabetic mice displayed increased food and water
intake with significantly increased urine output regardless of treatment (Table 2).
Diabetic mice had significant increases in HbA1c and blood glucose levels compared
with non-diabetic mice regardless of treatment (Table 2). Diabetes was associated
with increases in total cholesterol, triglycerides, HDL and LDL levels. All doses of
dh404 reduced these parameters although not to levels seen in the control mice with
the exception of the 20mg/kg/day treatment where triglycerides and LDL were not
affected by dh404.
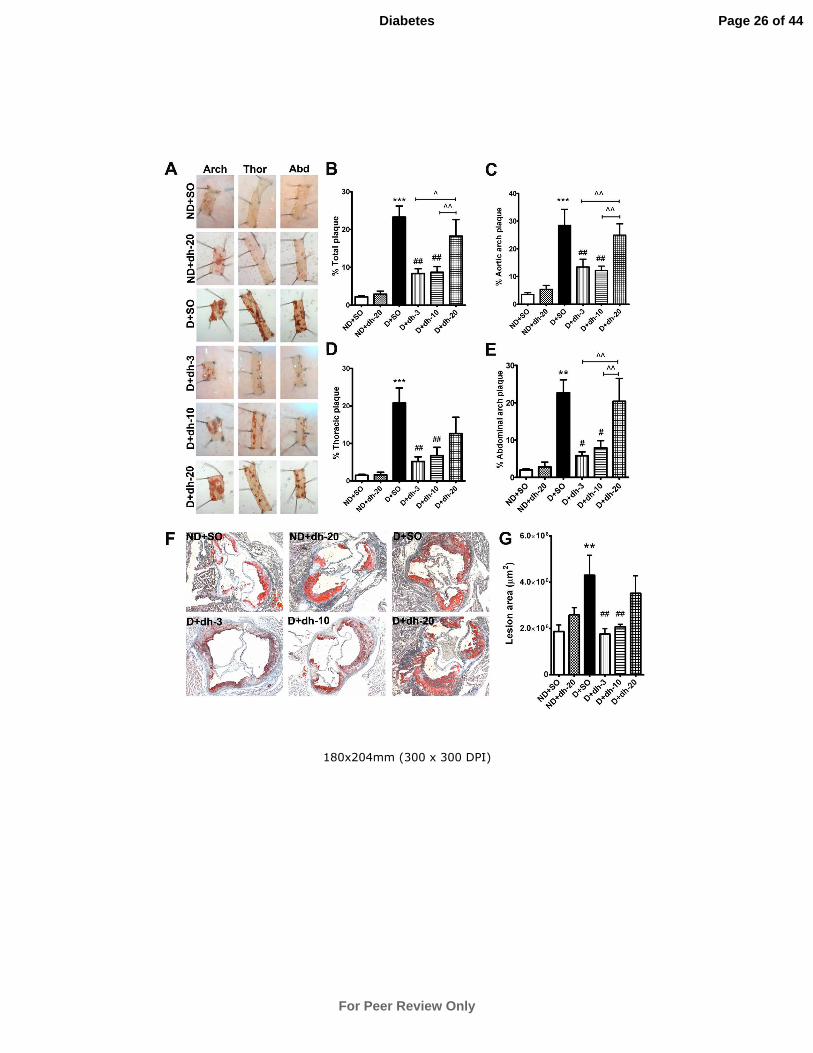
Atherosclerotic lesions
En face: Diabetes induced a significant increase in total plaque in ApoE-/- mice and
this increase was observed in the arch, thoracic and abdominal regions of the aorta.
Treatment of diabetic mice with dh404 at 3 and 10mg/kg/day significantly reduced
plaque within these regions. However, treatment with 20mg/kg/day dh404 failed to
reduce atherosclerotic plaque (Fig.1A-E).
Aortic sinus lesions: Diabetes significantly increased lesion deposition within the
aortic sinus after 20 weeks of diabetes (Fig.1F,G). This increase was significantly
attenuated after 18 weeks of dh404 treatment at 3 and 10mg/kg/day. Treatment with
20mg/kg/day dh404 failed to reduce diabetes-associated lesions.
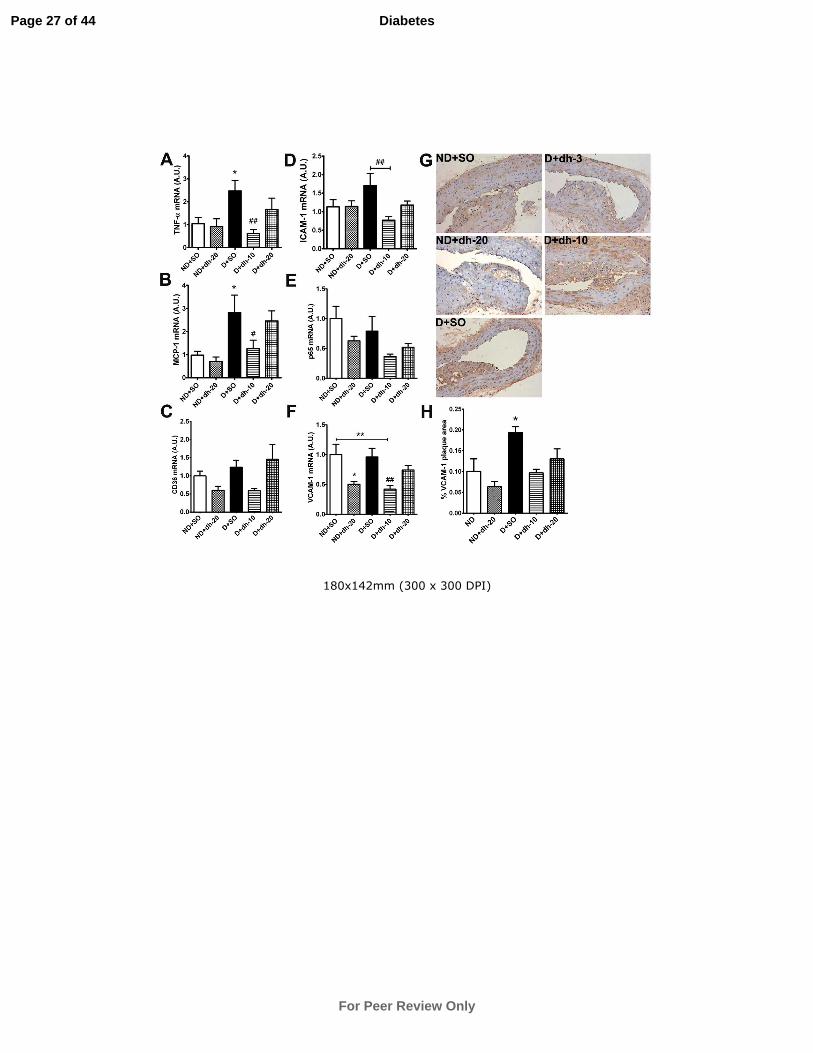
Adhesion and Inflammatory Gene and Protein Expression in Aorta
Gene expression of the inflammatory mediators TNF-α and MCP-1 was significantly
upregulated in diabetic aorta. This was significantly attenuated after 5 weeks of dh404
treatment at 10mg/kg/day but not at 20mg/kg/day (Fig.2A,B). No significant
Page 8 of 44
For Peer Review Only
Diabetes

9 | P a g e
differences were observed in CD36, ICAM-1, p65 and VCAM-1 gene expression
between vehicle-treated diabetic and non-diabetic mice (Fig.2C-F). However,
10mg/kg/day of dh404 reduced the expression of ICAM-1 and VCAM-1 when
compared with vehicle-treated counterparts. VCAM-1 protein levels were
significantly increased in diabetic plaque (P<0.05) and showed a trend towards a
reduction after 10 and 20mg/kg/day dh404 treatment (Fig.2G,H).
Oxidative Stress in Aortas
Analysis of 4-HNE, a marker of lipid hydroperoxides, within the vessel walls
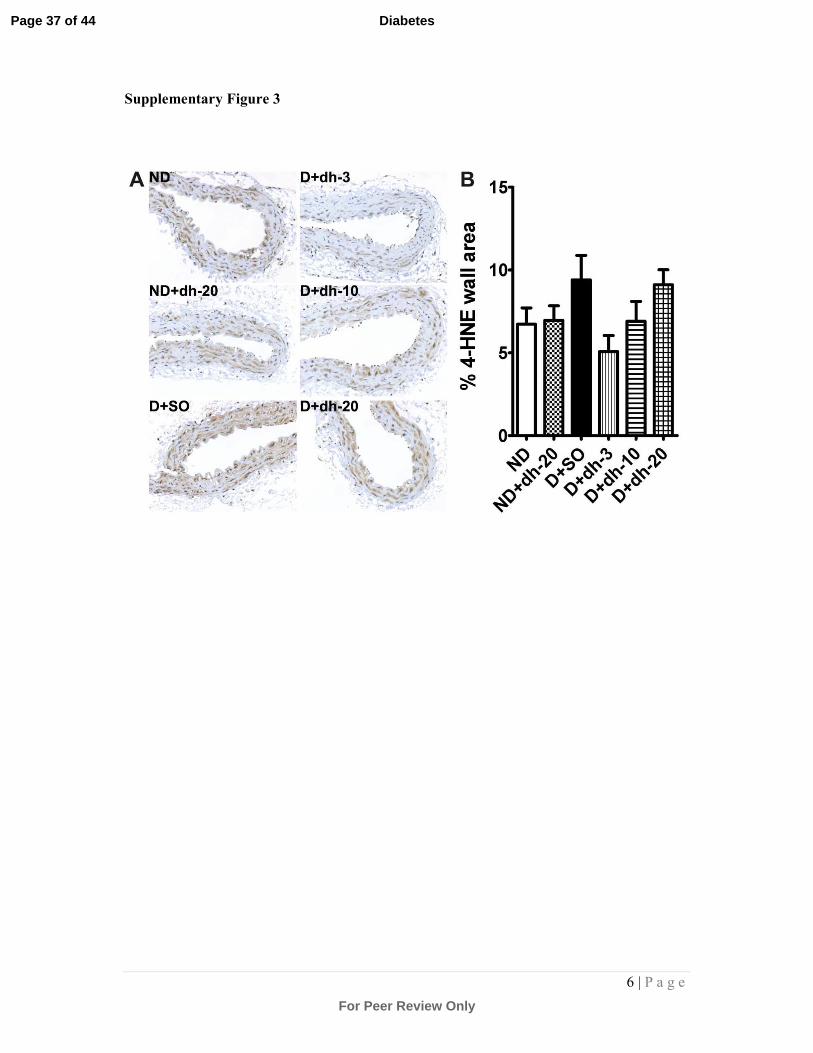
(Supplementary Fig.3) and plaques (Supplementary Fig.4) revealed a trend towards an
increase in diabetic mice. This was reduced after treatment with 3mg/kg/day dh404 in
the vessel wall, although this did not reach statistical significance. At the highest dose
of dh404, this trend was lost in vessel walls of the diabetic cohort. Plaque data
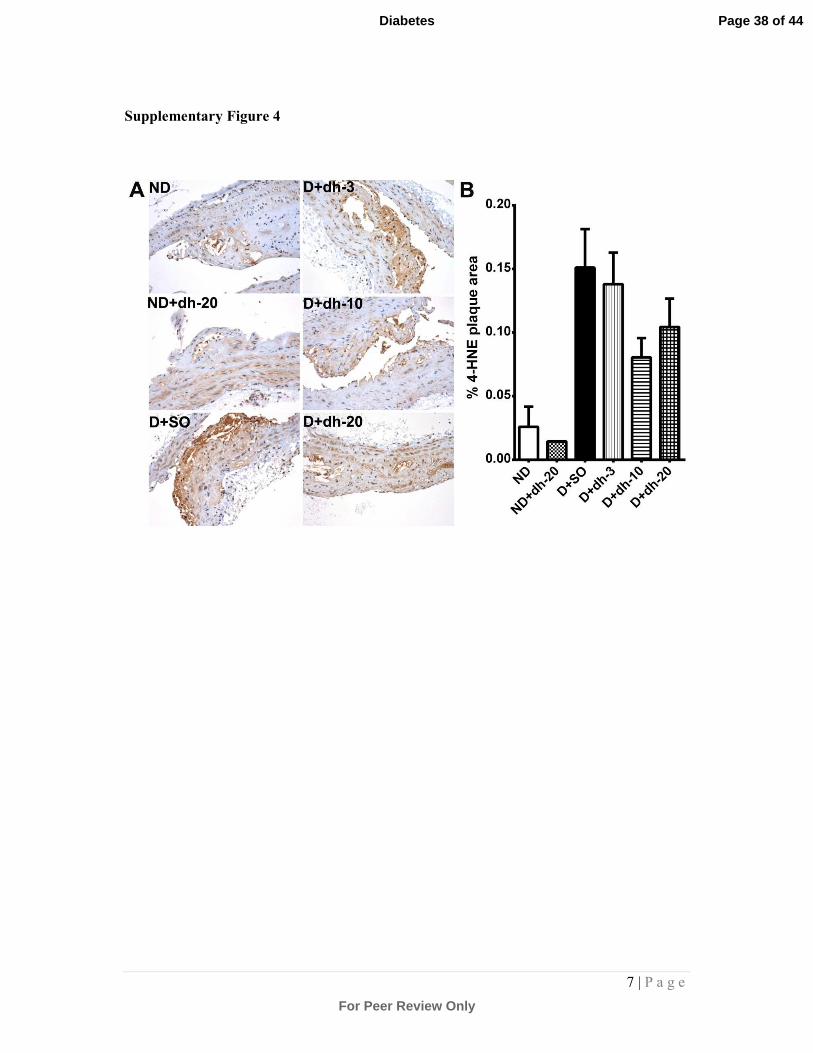
showed a trend towards a reduction in 4-HNE staining in diabetic mice treated with
10 and 20mg/kg/day dh404.
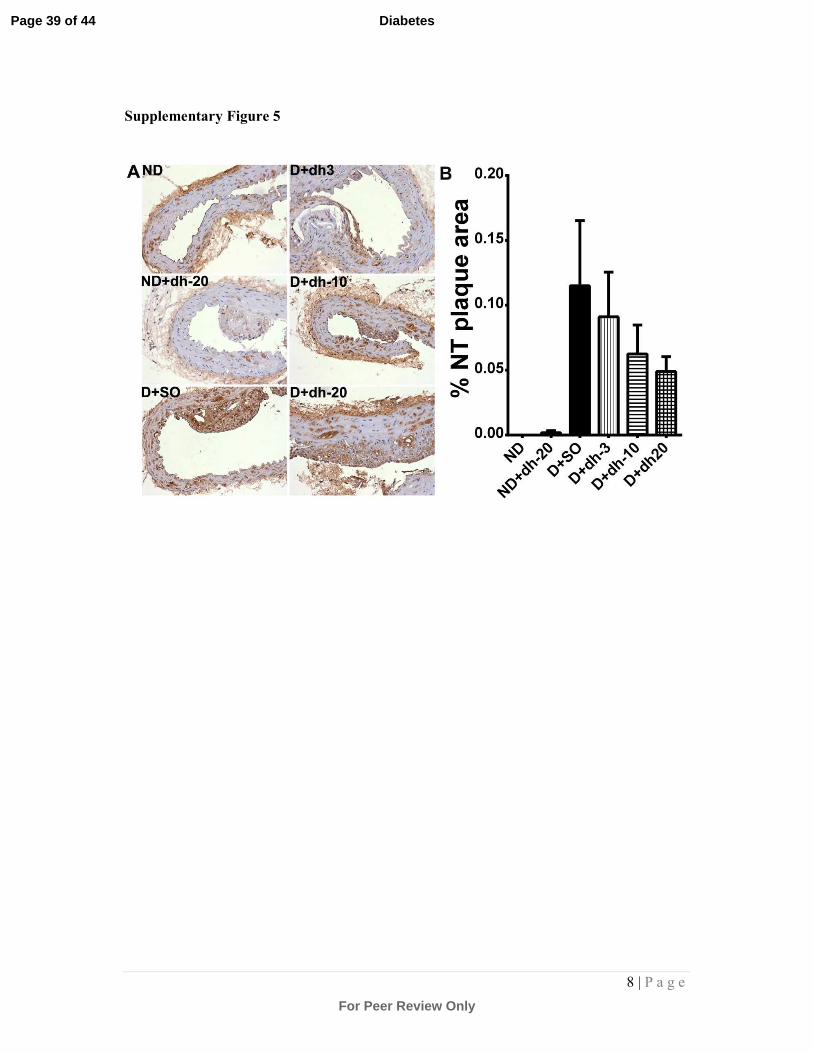
Analysis of nitrotyrosine, a marker of protein oxidative/nitrosative stress,
within plaque (Supplementary Fig.5) revealed a trend towards an increase in the
diabetic cohort, as observed previously by us in diabetic mice[27]. This trend was
lessened after treatment with dh404, with 20mg/kg/day showing the greatest reduction
in nitrotyrosine staining, albeit that this did not reach statistical significance.
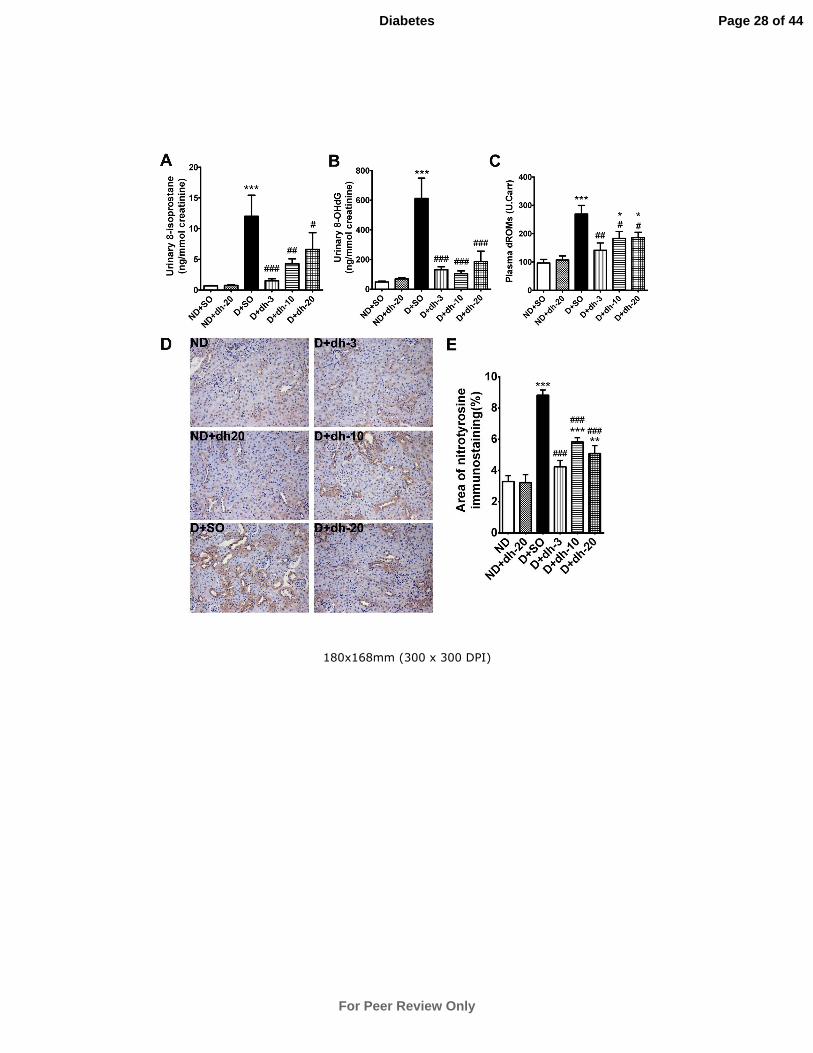
Oxidative Stress in urine and plasma
Urinary 8-isoprostane, 8-OHdG and plasma dROMs were all significantly upregulated
in vehicle-treated diabetic mice. Dh404 significantly attenuated these measures at all
doses tested (Fig.3A-C). Nitrotyrosine levels were assessed in the tubulointerstitial
region of the kidney and the diabetes-associated increase was significantly decreased
at all concentrations of dh404 (Fig.3D,E; P<0.001 vs non-diabetic controls).
Kidney Antioxidant and Inflammatory gene expression
The expression levels of Nrf2-responsive antioxidant genes in kidney cortex, after
5weeks of dh404 treatment, are shown in Fig.4A. NAD(P)H quinone oxidoreductase
(NQO1) was upregulated by diabetes and after treatment with 10 and 20mg/kg/day
Page 9 of 44
For Peer Review Only
Diabetes

10 | P a g e
dh404. Similarly, expression of GSH-S transferase was increased by diabetes but
further elevated after dh404 treatment. GPx1 expression increased in response to
dh404 treatment reaching significance at 20mg/kg/day dh404 in diabetic kidneys.
Inflammatory gene expression is shown in Fig.4B. Despite no change in IL-6
expression, MCP-1 gene expression was increased by diabetes and further increased
after 10 and 20mg/kg/day dh404, with significance reached at the highest dose of
dh404. The p65 subunit of NF-κB increased after dh404 treatment, reaching
significance at 20mg/kg/day dh404.
Renal Functional Parameters
Induction of diabetes led to a significant increase in the urinary albumin to creatinine
ratio which was significantly reduced by all doses of dh404.(Fig.4C). Elevations in
plasma cystatin C are an indication of declining glomerular filtration[30]. Plasma
cystatin C levels were significantly lower in vehicle-treated diabetic mice compared
to non-diabetic controls (Fig.4D) consistent with renal hyper-filtering[36]. Treatment
with dh404 did not alter plasma cystatin C levels, implying that dh404 did not worsen
renal function at the different doses.
Kidney Morphology
Glomerular injury: The percentage of PAS-positive material, indicative of mesangial
expansion, was significantly increased in diabetic glomeruli (Fig.4E,F). Furthermore,
the glomerulosclerosis index (GSI) of diabetic glomeruli was significantly higher than
that of non-diabetic controls (Fig.4G). Both mesangial expansion and GSI were
attenuated in diabetic mice treated with dh404 at 3mg/kg/day but this was not
observed at other doses of dh404.
Tubulointerstitial injury: TIA assessment revealed a significant increase in diabetic
kidneys which was significantly attenuated at all 3 doses of dh404 (Fig.4H) with the
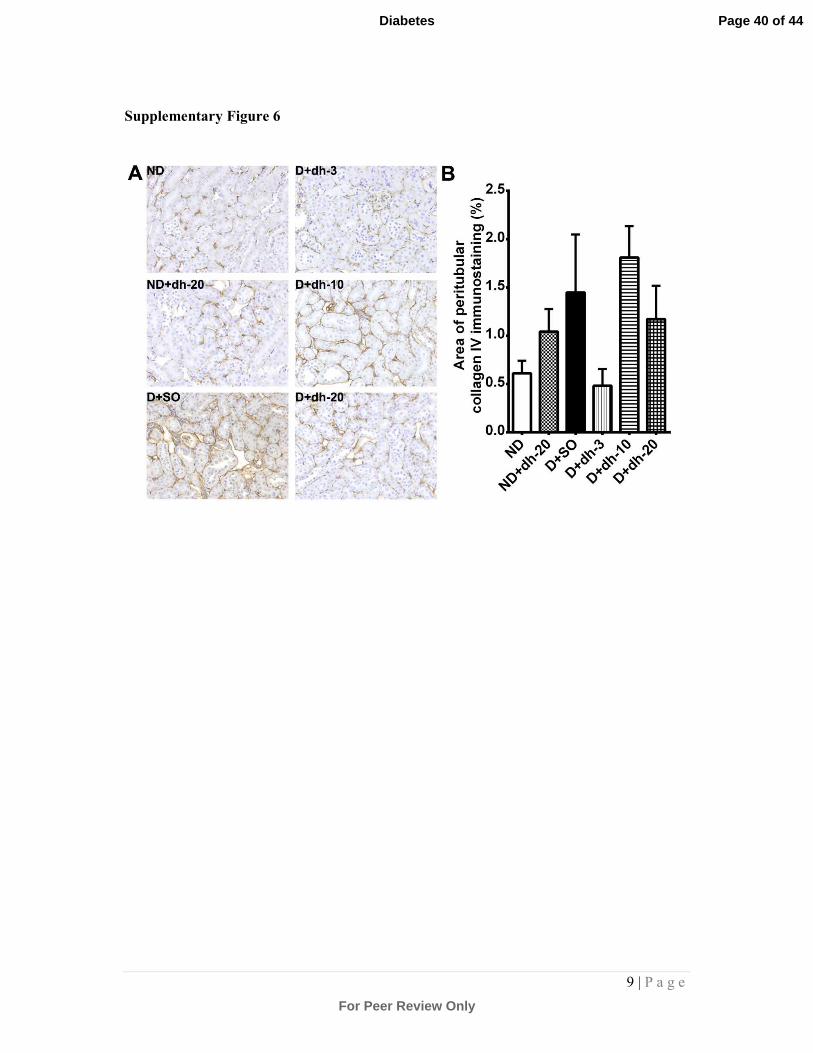
most significant reduction observed at 3mg/kg/day. Staining for collagen IV showed a
trend towards a reduction after 3mg/kg dh404 when compared to diabetic kidneys
treated with vehicle only (Supplementary Fig.6).
Page 10 of 44
For Peer Review Only
Diabetes

11 | P a g e
Liver function and pathology
Liver function was not affected by dh404 as plasma levels of ALT and AST were not
significantly different among treatment groups (Table 2). Histopathological
assessment of hepatocellular cytoplasmic rarification, an indicator of accumulated
glycogen, showed no significant differences after treatment with dh404, albeit that a
slight increase was observed in the non-diabetic (20mg/kg dh404) as well as diabetic
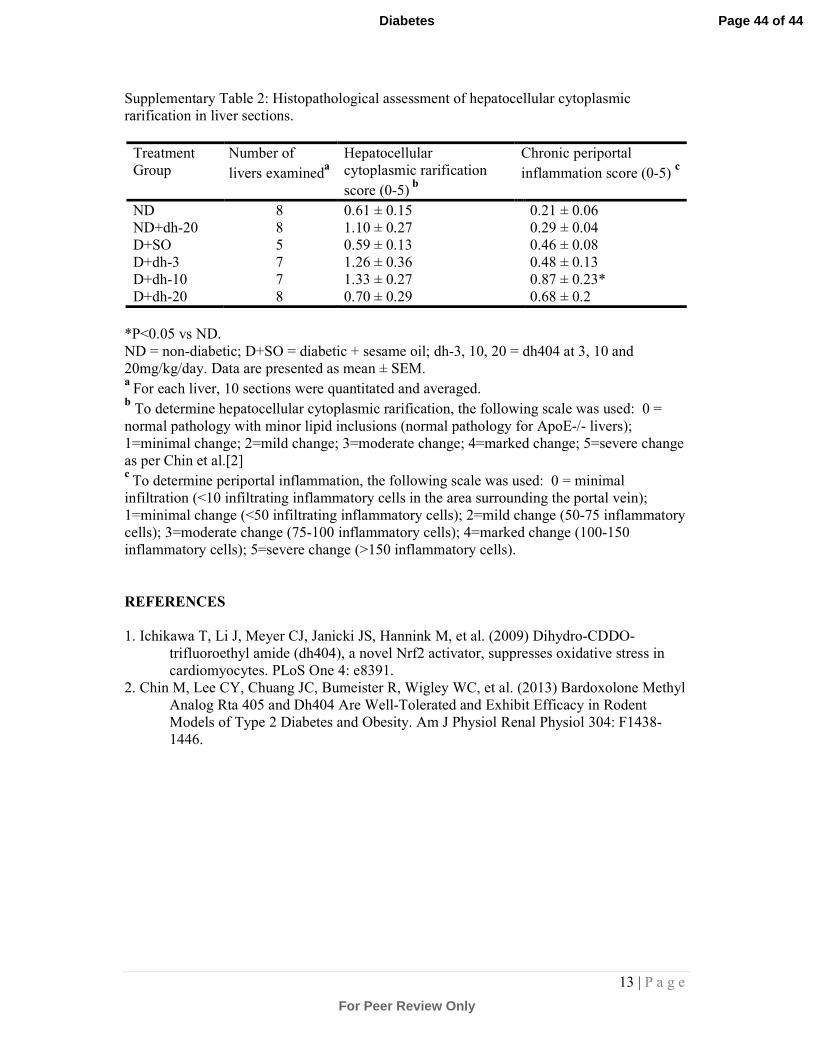
groups receiving 3 and 10mg/kg dh404 (Supplementary Table 2). Diabetes caused a
slight albeit non-significant increase in the number of infiltrating inflammatory cells.
The 10mg/kg dose of dh404 caused a statistically significant increase in infiltrating
inflammatory cells. However, this was considered mild with an infiltrate of less than
50 cells per portal vein. Thus, no significant treatment-related adverse liver pathology
was detected after 18 weeks of dh404 treatment (Supplementary Fig.5 7).
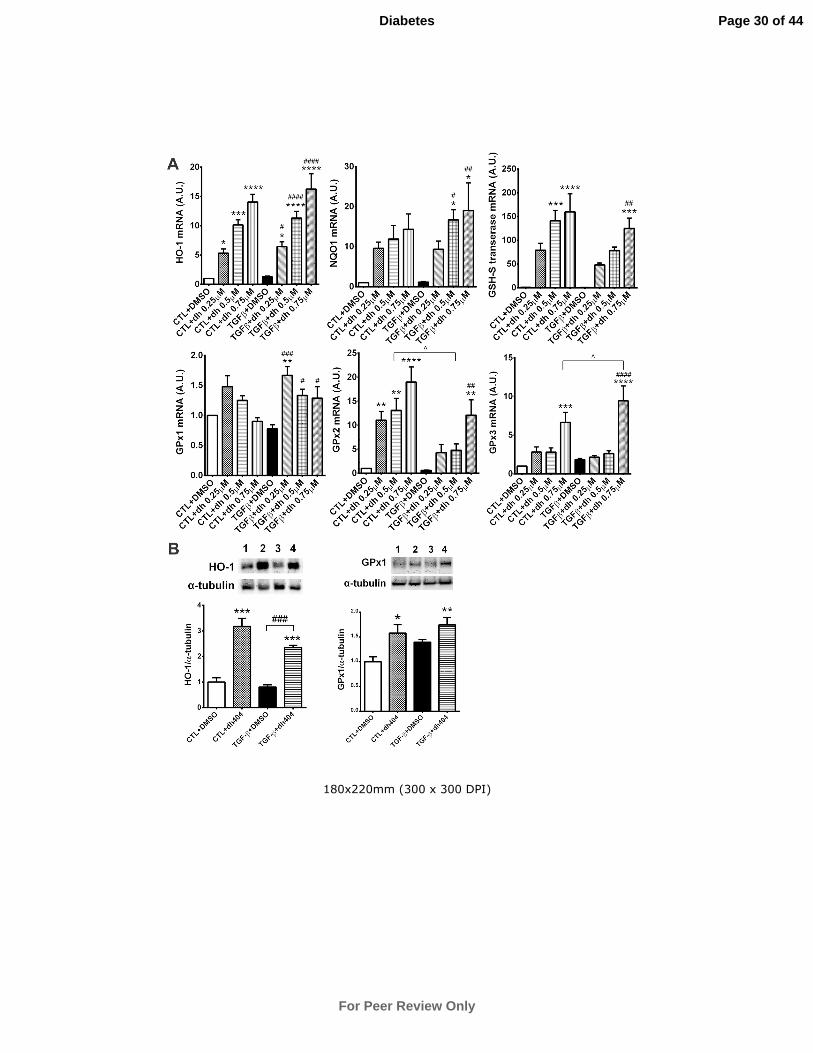
Antioxidant, inflammatory and fibrotic Gene and Protein Expression in NRK
cells
To complement our in vivo findings, we explored in vitro, in NRK cells in a dose-
dependent manner, the potential impact of dh404 on Nrf2-dependent antioxidant
genes, NQO1, heme oxygenase-1 (HO-1) and GSH-S transferase [37]. Gene
expression of these antioxidants was significantly upregulated by dh404, in the
presence or absence of TGF-β, with the greatest increase noted at the highest dose of
dh404 (Fig.5A). GPx1 gene expression was increased significantly by dh404 in the
presence of TGF-β. Dh404 increased GPx2 expression significantly in the absence of
TGF-β, and in the presence of TGF-β, this occurred at the highest dose of dh404.
GPx3 expression was increased significantly only at the highest dose of dh404 in the
presence or absence of TGF-β (Fig.5B). Changes in gene expression translated into
changes at the protein level as shown for HO-1 and GPx1 after 0.5µM dh404
treatment (Fig.5B)
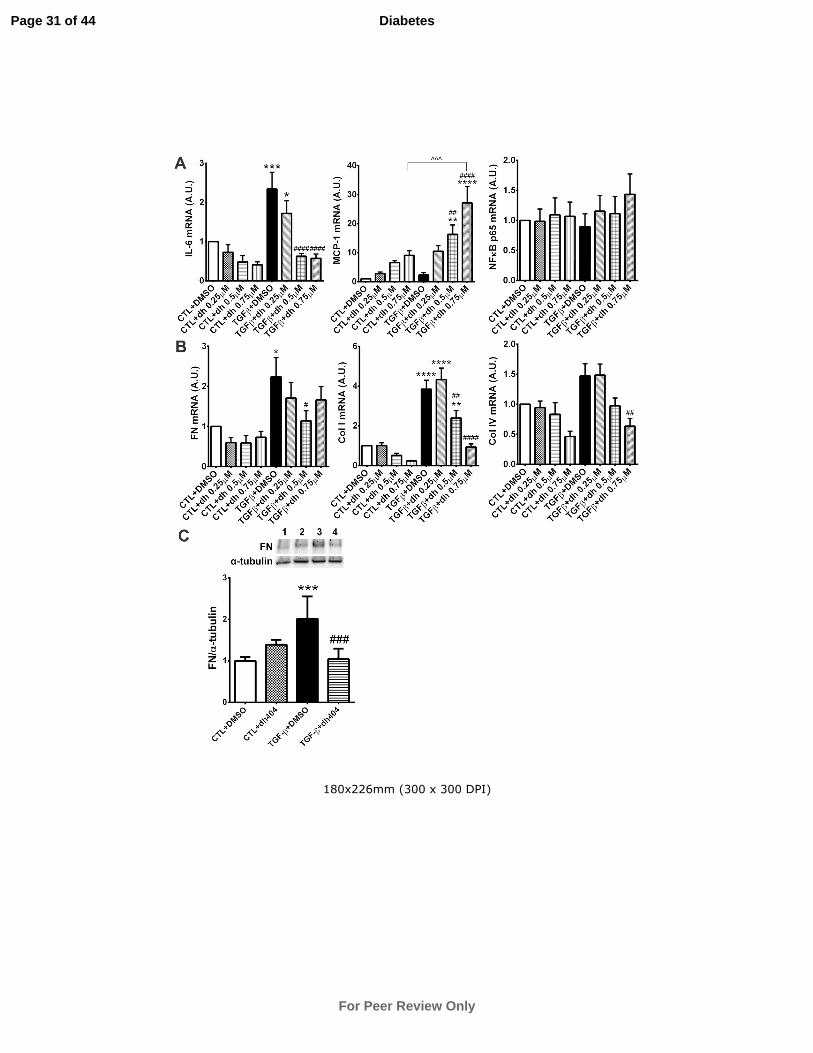
Despite positive reductions in IL-6 gene expression with increasing doses of
dh404 (Fig.6A), MCP-1 expression levels increased by ~20-fold at the highest dose of
dh404. p65 gene expression was unaffected by dh404 in TGF-β untreated cells but
showed a slight trend towards an increase at the highest dose of dh404 compared with
TGF-β treated controls (Fig.6A).
Page 11 of 44
For Peer Review Only
Diabetes

12 | P a g e
Treatment with TGF-β induced a significant increase in fibronectin and
collagen I and IV gene expression (Fig.6B). Importantly, dh404 significantly
attenuated the gene expression of these pro-fibrotic proteins, which was mirrored by a
similar protein profile for fibronectin. (Fig.6B,C).
Discussion
In the current study, the BM derivative, dh404, an Nrf2 agonist, has shown beneficial
effects in attenuating diabetes-associated atherosclerosis and nephropathy, albeit in an
inverse dose-related manner. To date, however, the role of Nrf2 in atherosclerosis
remains controversial. Studies have reported that Nrf2 may be pro-atherogenic using
ApoE/Nrf2 double knockout mice in which plaque and expression of the macrophage
scavenger receptor, CD36 were significantly reduced[38-40]. In contrast, the absence
of Nrf2 within the myeloid lineage in LDL receptor knockout (LDLR-/-) mice
aggravated both early and late stages of atherosclerosis[41,42]. Prior to the current
study, the role of Nrf2 modulation in diabetes-associated atherosclerosis remained an
unexplored area.
The results of this study show for the first time that dh404 lessens the
progression of diabetes-associated atherosclerosis. This was observed in all regions of
the aorta but appeared to be linked to the dose, with the lower doses of 3 and 10mg/kg
significantly lessening the progression of diabetic lesions, while the highest dose
(20mg/kg) failed to offer vascular protection. This was paralleled by similar
reductions in pro-inflammatory gene expression (TNF-α, MCP-1, ICAM-1 and
VCAM-1) at lower doses of dh404, while the 20mg/kg dose failed to afford such
protection. Interestingly, markers of oxidative stress such as urinary 8-isoprostane, 8-
OHdG and plasma dROMS were significantly attenuated by all concentrations of
dh404. Similarly, oxidative damage to proteins within plaque, as assessed by
nitrotyrosine staining, was reduced particularly at the highest dose of 20mg/kg dh404,
suggesting that at this dose there is no correlation between reductions in oxidative
stress and diabetes-associated atherosclerosis.
However, at lower doses, our results suggest that dh404 mediates its diabetes-
associated anti-atherogenic effects via its anti-inflammatory and antioxidative actions.
Triterpenoids have also been shown to regulate lipid metabolism[8,27]. Thus, it is
possible that the observed reductions are due in part to the action of dh404 on lipids
Page 12 of 44
For Peer Review Only
Diabetes

13 | P a g e
since we observed reductions in LDL, triglyceride and cholesterol after dh404
treatment, albeit that these reductions were rather modest. Additionally, the possible
minor improvement in metabolic control of diabetes by dh404 needs to be taken into
consideration. However, these rather modest changes are unlikely to have contributed
to the overall improvement in atherosclerosis. Furthermore, the lack of an effect on
atherosclerosis at the highest dose of dh404 could represent off-target effects of dh404
which may override any potential benefits of improved metabolic parameters in this
model.
In addition to these positive effects on the diabetic macrovasculature, we also
show significant improvements in renal function after 18 weeks of dh404 treatment.
This was reflected by improvements in the urinary albumin-to-creatinine ratio with
the lower doses of dh404 showing the greatest inhibition. It should be noted that the
finding of a reduction in urinary albumin-to-creatinine ratio contrasts with the human
data of the BEACON trial where despite improvements in eGFR, the urinary albumin-
to-creatinine ratio significantly increased in the BM-treated group.[22] A number of
possibilities could explain these discordant results such as a species-specific effect but
more likely relates to the timing of treatment. Indeed, human subjects had advanced
kidney disease (stage 4) prior to exposure to BM while in this study, mice were
treated soon after the establishment of diabetes as a preventative strategy. One of the
most prominent effects of BM in the BEAM[20] and BEACON[22] trials included an
increase in eGFR in subjects with renal impairment. Unfortunately most animal
models of DN including the STZ ApoE knockout mouse do not have declining GFR
but rather are models of hyperfiltration[36]. Indeed, in this study, analysis of plasma
cystatin C showed that diabetic mouse kidneys were hyperfiltering, a known early
phenomenon of kidney disease[30]. Thus, this model is unable to specifically test the
ability of Nrf2 agents such as dh404 to increase GFR.
Our structural analyses within the diabetic kidney showed significant
reductions in tubulointerstitial injury after treatment with dh404, with the lowest dose
providing the greatest protection. Similarly, only the lowest dose of dh404 lessened
the expression of the fibrosis marker, collagen IV, within this region of the kidney
albeit that these reductions fell just short of significance. This was also accompanied
by significant reductions in oxidative stress within the tubular region of the kidney
after dh404 treatment. Additionally, we also observed significant improvements in
Page 13 of 44
For Peer Review Only
Diabetes

14 | P a g e
glomerulosclerosis at the lowest dose of dh404. Our results therefore contrast with the
study of Zoja et al.[23], where it was reported that dh404 failed to show beneficial
effects on proteinuria and significantly worsened tubular damage in the diabetic
Zucker fat rat. Our study also contrasts with their study in that we did not observe any
adverse effects of drug treatment on liver structure and function, and failed to detect
any renal pseudotumors. Differences in outcome between our study and that of Zoja et
al. may reflect subtle disparity in drug dosage as well as differences in drug
bioavailability between species. Our findings are however, more in agreement with
Chin et al.[19] where it was shown that the BM analogues, RTA 405 and dh404, were
well tolerated in various rodent models of Type 2 diabetes. Our study is also in
agreement with the notion that activators of Nrf2 protect against diabetic
nephropathy[9].
It is of interest that in the present study, the lower doses of dh404 appear to
limit end-organ injury whereas this benefit is lost at higher dosage. Therefore, it is
critical to determine whether the higher dose of drug did not work in vivo due to a
lack of an increase in antioxidant production or due to an off-target effect. To address
this issue, we analysed several Nrf2-responsive antioxidant genes (NQO1, GSH-S
transferase and GPx1) in kidney cortex. Indeed, the gene expression of NQO1 and
GSH-S transferase are further increased in response to 20mg/kg dh404. In addition,
cell culture experiments confirmed the responsiveness of Nrf2 genes to dh404 in a
dose-dependent manner. Taken together, our results indicate that a lack of end-organ
protection by the highest dose of dh404 cannot be explained by a lack of antioxidant
responsiveness to dh404.
Possible off-target effects of the drug that may account for the loss of
protection at higher doses were assessed. Indeed, pro-inflammatory mediators such as
MCP-1 were significantly upregulated in a dose-responsive manner with the highest
dose of dh404 resulting in an ~20-fold increase in MCP-1 gene expression in NRK
cells. Interestingly, the highest dose of dh404 also caused a significant increase in
MCP-1 expression in the kidneys of dh404-treated mice. The latter finding was also
accompanied by a significant increase in the expression of the p65 subunit of the pro-
inflammatory transcription factor, NF-κB, in the high-dose treated diabetic kidney.
Similarly, higher aortic MCP-1 expression correlated with greater plaque. Thus, our
data suggest that dh404 might act to promote inflammation at higher doses.
Page 14 of 44
For Peer Review Only
Diabetes

15 | P a g e
Our in vitro results suggest that dh404 regulates its anti-fibrotic effects in the
kidney, at least partly, through the inhibition of TGF-β activity, since both TGF-β-
stimulated fibronectin and collagen I and IV expression were inhibited by dh404 in rat
kidney tubular cells. Our results are therefore consistent with previous studies that
demonstrate Nrf2-mediated renal protection through the inhibition of TGF-β promoter
activity and its downstream pathways[9,43].
Finally, these findings need to be considered in the clinical context. The
systemic exposure of dh404 at the lower doses in this study, corrected for the potency
difference between dh404 and BM, was similar to that observed in both the BEAM
and BEACON studies. Therefore, protection from murine end-organ injury occurs at
clinically relevant exposures, whereas this beneficial effect was lost at higher doses.
Thus, as seen in the Nrf2 knockout studies[38-40], there may be a therapeutic window
for this drug class and detailed preclinical and subsequently clinical studies with such
agents will need to be tested in a careful dose-dependent manner.
In summary, our data have shown inverse dose-dependent improvements in
diabetes-associated atherosclerosis and diabetic nephropathy through the use of an
Nrf2 activator, dh404, via reductions in pro-inflammatory mediators and oxidative
stress in our pre-clinical model of STZ-induced diabetes. The anti-atherogenic
function of AIM compounds, and dh404 in particular, have not previously been
explored. Our study therefore potentially extends the pharmacotherapeutic benefits of
this class of compound to the protection against DAA, a major complication affecting
diabetic patients. Our study also raises the possibility that dh404 functions to lessen
both diabetic kidney injury and diabetes-associated atherosclerosis, a highly desirable
clinical outcome since cardiovascular and renal disease often occur concurrently[44].
Finally, our findings, showing greater efficacy with respect to both cardiovascular and
renal outcomes at lower dh404 doses, highlights the importance of determining the
effective pharmacokinetic range for this class of drug. Further preclinical
characterisation of AIM compounds is therefore vital before one can assess the true
potential of this drug class in the clinic.
Page 15 of 44
For Peer Review Only
Diabetes

16 | P a g e
Acknowledgements
S.M.T. is supported by a JDRF International Postdoctoral Fellowship. We would like
to acknowledge Katherine Ververis for her excellent technical assistance with the
NRK cell studies.
A.S., N.S., T.K and D.Y.C.Y researched data. J.B.d.H. and S.M.T. wrote the
manuscript, researched data and analysed results. C.M., K.W.W. and M.E.C
reviewed/edited the manuscript. Judy B. de Haan is the guarantor of this work and, as
such, had full access to all the data in the study and takes responsibility for the
integrity of the data and the accuracy of the data analysis.
SMT, AS, NS, DYCY, MEC, T.K and JBdH report grants and non-financial support
from Reata Pharmaceuticals Inc. during the conduct of the study.
KWW and CM are Employees of Reata Pharmaceuticals Inc. and report personal fees
from Reata Pharmaceuticals during the conduct of the study.
Page 16 of 44
For Peer Review Only
Diabetes

17 | P a g e
References
1. Haffner SM, Lehto S, Ronnemaa T, Pyorala K, Laakso M (1998) Mortality from
coronary heart disease in subjects with type 2 diabetes and in nondiabetic
subjects with and without prior myocardial infarction. N Engl J Med 339: 229-
234.
2. Reutens AT (2013) Epidemiology of diabetic kidney disease. Med Clin North Am
97: 1-18.
3. Mittal M, Siddiqui MR, Tran K, Pothireddy S, Malik AB (2013) Reactive Oxygen
Species in Inflammation and Tissue Injury. Antioxid Redox Signal.
4. de Haan JB, Cooper ME (2011) Targeted antioxidant therapies in hyperglycemia-
mediated endothelial dysfunction. Front Biosci (Schol Ed) 3: 709-729.
5. Jung KA, Kwak MK (2010) The Nrf2 system as a potential target for the
development of indirect antioxidants. Molecules 15: 7266-7291.
6. de Haan JB (2011) Nrf2 activators as attractive therapeutics for diabetic
nephropathy. Diabetes 60: 2683-2684.
7. Li B, Liu S, Miao L, Cai L (2012) Prevention of diabetic complications by activation
of Nrf2: diabetic cardiomyopathy and nephropathy. Exp Diabetes Res 2012:
216512.
8. Ruiz S, Pergola PE, Zager RA, Vaziri ND (2013) Targeting the transcription factor
Nrf2 to ameliorate oxidative stress and inflammation in chronic kidney
disease. Kidney Int 83: 1029-1041.
9. Zheng H, Whitman SA, Wu W, Wondrak GT, Wong PK, et al. (2011) Therapeutic
potential of Nrf2 activators in streptozotocin-induced diabetic nephropathy.
Diabetes 60: 3055-3066.
10. Kobayashi M, Li L, Iwamoto N, Nakajima-Takagi Y, Kaneko H, et al. (2009) The
antioxidant defense system Keap1-Nrf2 comprises a multiple sensing
mechanism for responding to a wide range of chemical compounds. Mol Cell
Biol 29: 493-502.
11. Lee Y, Shin DH, Kim JH, Hong S, Choi D, et al. (2010) Caffeic acid phenethyl ester-
mediated Nrf2 activation and IkappaB kinase inhibition are involved in
NFkappaB inhibitory effect: structural analysis for NFkappaB inhibition. Eur J
Pharmacol 643: 21-28.
12. Dinkova-Kostova AT, Liby KT, Stephenson KK, Holtzclaw WD, Gao X, et al. (2005)
Extremely potent triterpenoid inducers of the phase 2 response: correlations
of protection against oxidant and inflammatory stress. Proc Natl Acad Sci U S
A 102: 4584-4589.
13. Liby KT, Sporn MB (2012) Synthetic oleanane triterpenoids: multifunctional drugs
with a broad range of applications for prevention and treatment of chronic
disease. Pharmacol Rev 64: 972-1003.
14. Aleksunes LM, Goedken MJ, Rockwell CE, Thomale J, Manautou JE, et al. (2010)
Transcriptional regulation of renal cytoprotective genes by Nrf2 and its
potential use as a therapeutic target to mitigate cisplatin-induced
nephrotoxicity. J Pharmacol Exp Ther 335: 2-12.
15. Tanaka Y, Aleksunes LM, Goedken MJ, Chen C, Reisman SA, et al. (2008)
Coordinated induction of Nrf2 target genes protects against iron
Page 17 of 44
For Peer Review Only
Diabetes

18 | P a g e
nitrilotriacetate (FeNTA)-induced nephrotoxicity. Toxicol Appl Pharmacol 231:
364-373.
16. Wu QQ, Wang Y, Senitko M, Meyer C, Wigley WC, et al. (2011) Bardoxolone
methyl (BARD) ameliorates ischemic AKI and increases expression of
protective genes Nrf2, PPARgamma, and HO-1. Am J Physiol Renal Physiol
300: F1180-1192.
17. Ungvari ZI, Bailey-Downs L, Gautam T, Jimenez R, Losonczy G, et al. (2011)
Adaptive induction of NF-E2-Related Factor-2-driven antioxidant genes in
endothelial cells in response to hyperglycemia. Am J Physiol Heart Circ
Physiol 300: H1133-1140.
18. Xue M, Qian Q, Adaikalakoteswari A, Rabbani N, Babaei-Jadidi R, et al. (2008)
Activation of NF-E2-related factor-2 reverses biochemical dysfunction of
endothelial cells induced by hyperglycemia linked to vascular disease.
Diabetes 57: 2809-2817.
19. Pergola PE, Krauth M, Huff JW, Ferguson DA, Ruiz S, et al. (2011) Effect of
bardoxolone methyl on kidney function in patients with T2D and Stage 3b-4
CKD. Am J Nephrol 33: 469-476.
20. Pergola PE, Raskin P, Toto RD, Meyer CJ, Huff JW, et al. (2011) Bardoxolone
methyl and kidney function in CKD with type 2 diabetes. N Engl J Med 365:
327-336.
21. de Zeeuw D, Akizawa T, Agarwal R, Audhya P, Bakris GL, et al. (2013) Rationale
and trial design of Bardoxolone Methyl Evaluation in Patients with Chronic
Kidney Disease and Type 2 Diabetes: the Occurrence of Renal Events
(BEACON). Am J Nephrol 37: 212-222.
22. de Zeeuw D AT, Audhya P, Bakris GL, Chin M, Christ-Schmidt H, Goldsberry A,
Houser M, Krauth M, Lambers Heerspink HJ, McMurray JJ, Meyer CJ, Parving
H-H, Remuzzi G, Toto RD, Vaziri ND, Wanner C, Wittes J, Wrolstad D, Chertow
GM. (2013) Bardoxolone Methyl in Type 2 diabetes and Stage 4 chronic
kidney disease. NEJM.
23. Zoja C, Corna D, Nava V, Locatelli M, Abbate M, et al. (2013) Analogs of
bardoxolone methyl worsen diabetic nephropathy in rats with additional
adverse effects. Am J Physiol Renal Physiol 304: F808-819.
24. Chin M, Lee CY, Chuang JC, Bumeister R, Wigley WC, et al. (2013) Bardoxolone
Methyl Analog Rta 405 and Dh404 Are Well-Tolerated and Exhibit Efficacy in
Rodent Models of Type 2 Diabetes and Obesity. Am J Physiol Renal Physiol
304: F1438-1446.
25. Ichikawa T, Li J, Meyer CJ, Janicki JS, Hannink M, et al. (2009) Dihydro-CDDO-
trifluoroethyl amide (dh404), a novel Nrf2 activator, suppresses oxidative
stress in cardiomyocytes. PLoS One 4: e8391.
26. Lewis P, Stefanovic N, Pete J, Calkin AC, Giunti S, et al. (2007) Lack of the
antioxidant enzyme glutathione peroxidase-1 accelerates atherosclerosis in
diabetic apolipoprotein E-deficient mice. Circulation 115: 2178-2187.
27. Chew P, Yuen DY, Koh P, Stefanovic N, Febbraio MA, et al. (2009) Site-specific
antiatherogenic effect of the antioxidant ebselen in the diabetic
apolipoprotein E-deficient mouse. Arterioscler Thromb Vasc Biol 29: 823-830.
Page 18 of 44
For Peer Review Only
Diabetes

19 | P a g e
28. Chew P, Yuen DY, Stefanovic N, Pete J, Coughlan MT, et al. (2010)
Antiatherosclerotic and renoprotective effects of ebselen in the diabetic
apolipoprotein E/GPx1-double knockout mouse. Diabetes 59: 3198-3207.
29. Krege JH, Hodgin JB, Hagaman JR, Smithies O (1995) A noninvasive computerized
tail-cuff system for measuring blood pressure in mice. Hypertension 25:
1111-1115.
30. Grubb A (1992) Diagnostic value of analysis of cystatin C and protein HC in
biological fluids. Clin Nephrol 38 Suppl 1: S20-27.
31. Cornelli U, Terranova R, Luca S, Cornelli M, Alberti A (2001) Bioavailability and
Antioxidant Activity of Some Food Supplements in Men and Women Using
the D-Roms Test as a Marker of Oxidative Stress. The Journal of Nutrition
131: 3208-3211.
32. Cornelli U, Terranova R, Luca S, Cornelli M, Alberti A (2001) Bioavailability and
antioxidant activity of some food supplements in men and women using the
D-Roms test as a marker of oxidative stress. J Nutr 131: 3208-3211.
33. Forbes JM, Thallas V, Thomas MC, Founds HW, Burns WC, et al. (2003) The
breakdown of preexisting advanced glycation end products is associated with
reduced renal fibrosis in experimental diabetes. FASEB J 17: 1762-1764.
34. Tan SM, Stefanovic N, Tan G, Wilkinson-Berka JL, de Haan JB (2013) Lack of the
antioxidant glutathione peroxidase-1 (GPx1) exacerbates retinopathy of
prematurity in mice. Invest Ophthalmol Vis Sci 54: 555-562.
35. Herman-Edelstein M, Thomas MC, Thallas-Bonke V, Saleem M, Cooper ME, et al.
(2011) Dedifferentiation of immortalized human podocytes in response to
transforming growth factor-beta: a model for diabetic podocytopathy.
Diabetes 60: 1779-1788.
36. Ott IM, Alter ML, von Websky K, Kretschmer A, Tsuprykov O, et al. (2012) Effects
of stimulation of soluble guanylate cyclase on diabetic nephropathy in
diabetic eNOS knockout mice on top of angiotensin II receptor blockade. PLoS
One 7: e42623.
37. Ohtsuji M, Katsuoka F, Kobayashi A, Aburatani H, Hayes JD, et al. (2008) Nrf1 and
Nrf2 play distinct roles in activation of antioxidant response element-
dependent genes. J Biol Chem 283: 33554-33562.
38. Harada N, Ito K, Hosoya T, Mimura J, Maruyama A, et al. (2012) Nrf2 in bone
marrow-derived cells positively contributes to the advanced stage of
atherosclerotic plaque formation. Free Radic Biol Med 53: 2256-2262.
39. Barajas B, Che N, Yin F, Rowshanrad A, Orozco LD, et al. (2011) NF-E2-related
factor 2 promotes atherosclerosis by effects on plasma lipoproteins and
cholesterol transport that overshadow antioxidant protection. Arterioscler
Thromb Vasc Biol 31: 58-66.
40. Sussan TE, Jun J, Thimmulappa R, Bedja D, Antero M, et al. (2008) Disruption of
Nrf2, a key inducer of antioxidant defenses, attenuates ApoE-mediated
atherosclerosis in mice. PLoS ONE 3: e3791.
41. Collins AR, Gupte AA, Ji R, Ramirez MR, Minze LJ, et al. (2012) Myeloid deletion of
nuclear factor erythroid 2-related factor 2 increases atherosclerosis and liver
injury. Arterioscler Thromb Vasc Biol 32: 2839-2846.
Page 19 of 44
For Peer Review Only
Diabetes

20 | P a g e
42. Ruotsalainen AK, Inkala M, Partanen ME, Lappalainen JP, Kansanen E, et al.
(2013) The absence of macrophage Nrf2 promotes early atherogenesis.
Cardiovasc Res 98: 107-115.
43. Jiang T, Huang Z, Lin Y, Zhang Z, Fang D, et al. (2010) The protective role of Nrf2
in streptozotocin-induced diabetic nephropathy. Diabetes 59: 850-860.
44. Tonelli M, Muntner P, Lloyd A, Manns BJ, Klarenbach S, et al. (2012) Risk of
coronary events in people with chronic kidney disease compared with those
with diabetes: a population-level cohort study. Lancet 380: 807-814.
Page 20 of 44
For Peer Review Only
Diabetes

21 | P a g e
Figure legends
Figure 1: Diabetes-associated lesion formation in the aorta and aortic sinus is
attenuated by dh404 after 18 weeks of treatment. Aortas were stained with Sudan
IV-Herxheimer’s solution and plaques were stained red (A). Treatment with dh404 at
3 and 10mg/kg/day resulted in an attenuation of total plaque formation in the aortas of
diabetic mice (B), and similar effects were found in the aortic arch (C), thoracic (D)
and abdominal regions (E). Cryosections of aortic sinus were stained with Oil Red O
to detect plaque formation after 18 weeks of dh404 treatment (F). At 3 and 10
mg/kg/day, plaque formation was significantly attenuated in diabetic mice when
compared to their vehicle-treated counterparts (G). Data are mean ± SEM. **P<0.01,
***P<0.001 vs ND+SO; #P<0.05,
##P<0.01 vs D+SO;
^P<0.05,
^^P<0.01 vs as
indicated. ND = non-diabetic mice; SO = sesame oil; dh-3,-10,-20 = dh404 at 3, 10 or
20mg/kg/day; D = diabetic mice.
Figure 2: Adhesion and inflammatory markers in aorta are reduced by dh404.
The gene expression of adhesion and inflammatory markers in the aorta was assessed
by qRT-PCR after 5 weeks of treatment (A-F). VCAM-1 protein expression was
examined using immunohistochemistry in aortic plaque (G) and the quantitation is
shown in (H). Data are mean ± SEM. *P<0.05, **P<0.01 vs ND+SO; #P<0.05,
##P<0.01 vs D+SO. TNF-α = tumour necrosis factor-α; MCP-1 = monocyte
chemotactic protein-1; ICAM-1 = intercellular adhesion molecule-1; VCAM-1;
vascular cell adhesion molecule-1; ND = non-diabetic mice; SO = sesame oil; dh-3,-
10,-20 = dh404 at 3, 10 or 20mg/kg/day; D = diabetic mice. For qRT-PCR analysis,
n=6-10 (MCP-1), 7-10 (TNF-α), and 8-10 (CD36, ICAM1, p65 and VCAM-1)
aortas/group.
Figure 3: Diabetes-associated oxidative stress is attenuated by dh404 after 18
weeks of treatment. Oxidative stress was assessed by urine levels of 8-isoprostane
(A) and 8-OHdG (B) and plasma levels of dROMs (C). All of these markers were
attenuated in the diabetic mice by dh404. Nitrotyrosine immunostaining revealed that
diabetes was associated with an increase in nitrotyrosine protein expression and this
was attenuated by dh404 at all doses (D and E). Data are mean ± SEM. *P<0.05,
Page 21 of 44
For Peer Review Only
Diabetes

22 | P a g e
**P<0.01, ***P<0.001 vs ND+SO; #P<0.05,
##P<0.01,
###P<0.001 vs D+SO. ND =
non-diabetic mice; SO = sesame oil; dh-3,-10,-20 = dh404 at 3, 10 or 20mg/kg/day; D
= diabetic mice.
Figure 4: Dh404 increases antioxidant, as well as inflammatory gene expression
after 5 weeks of treatment, and improves kidney function and attenuates
glomerulosclerosis and tubulointerstitial injury after 18 weeks of treatment. The
gene expression of antioxidants, NQO1, GSH-S transferase and GPx1, were increased
in the kidney cortex by dh404 after 5 weeks of treatment (A). However, inflammatory
genes such as MCP-1 and NKκB were also upregulated by dh404 (B). Diabetes-
associated increase in urinary albumin to creatinine ratio (UACR) was reduced by
dh404 (C), however dh404 treatment had no effect on plasma cystatin C levels (D).
Glomerulosclerosis was assessed by measuring Periodic acid-Schiff (PAS)-stained
area per glomerulus (indicative of mesangial expansion) and also scored for
glomerulosclerosis index (GSI). Photomicrographs of representative glomeruli are
shown in (E). Treatment of dh404 at 3mg/kg/day for 18 weeks significantly
attenuated mesangial expansion (F) and GSI (G). The percentage tubulointerstitial
injury was determined from PAS-stained sections by point-counting and is shown in
(H). Data are mean ± SEM. *P<0.05, **P<0.01, ***P<0.001 vs ND+SO; #P<0.05,
##P<0.01,
###P<0.001 vs D+SO. ND = non-diabetic mice; SO = sesame oil; dh-3,-10,-
20 = dh404 at 3, 10 or 20mg/kg/day; D = diabetic mice.
Page 22 of 44
For Peer Review Only
Diabetes

23 | P a g e
Figure 5: In vitro analysis of Nrf2-responsive antioxidant genes in NRK cells
after dh404 treatment. NRK cells were treated with either DMSO, dh404, TGF-
β+DMSO or TGF-β+dh404 for 72 hours as detailed in the methods. Gene expression
profiles of hemeoxygenase-1 (HO1), NAD(P)H dehydrogenase (quinone 1) (NQO1),
GSH-S transferase, glutathione peroxidase (GPx)1, GPx2 and GPx3 are shown in A.
Dh404, in most cases, caused dose-dependent increases in the gene expression of
most antioxidants investigated either in the presence or absence of TGF-β treatment.
Protein levels for HO-1 and GPx1 after treatment of NRK cells with 0.5µM dh404 for
72hr are shown in B. Data are mean ± SEM from 5 separate experiments for qPCR
analyses (n=5) and 3 separate experiments for Western analysis (n=3). For the qPCR
studies, data was normalized to controls (given arbitrary value =1) as fold change and
then averaged over replicate 5 experiments. *P<0.05, **P<0.01, ***P<0.001,
****P<0.0001 vs CTL+DMSO; ##
P<0.01, ###
P<0.001, ####
P<0.0001 vs TGF-
β+DMSO; ^P<0.05 CTL+dh404 vs TGF-β+dh404. CTL = control; DMSO = dimethyl
sulfoxide. Lane 1 = DMSO; lane 2 = dh404; lane 3 = TGF-β+DMSO; lane 4 = TGF-
β+dh404.
Figure 6: In vitro analysis of Nrf2-responsive anti-inflammatory and fibrosis
genes in NRK cells after dh404 treatment. NRK cells were treated with either
DMSO, dh404, TGF-β+DMSO or TGF-β+dh404 for 72 hours as detailed in the
methods. Gene expression profiles of IL-6, MCP-1 and the p65 subunit of NF-κB are
shown in A. Gene expression of fibronectin (FN), collagen I and IV are shown in B.
Despite decreases in IL-6 expression with increasing doses of dh404, MCP-1 showed
a significant increase in gene expression at higher doses of dh404. Collagen I and IV
as well as FN gene expression was significantly attenuated by dh404. FN protein
levels, shown in C, were significantly reduced after 0.5µM dh404 treatment. Data are
mean ± SEM from 5 separate experiments for qPCR analyses (n=5) and 3 separate
experiments for Western analysis (n=3). For the qPCR studies, data was normalized
to controls (given arbitrary value =1) as fold change and then averaged over replicate
5 experiments. *P<0.05, **P<0.01, ***P<0.001, ****P<0.0001 vs CTL+DMSO;
##P<0.01,
###P<0.001,
####P<0.0001 vs TGF-β+DMSO; ^^^P<0.001 CTL+dh404 vs
TGF-β+dh404. CTL = control; DMSO = dimethyl sulfoxide. Lane 1 = DMSO; lane 2
= dh404; lane 3 = TGF-β+DMSO; lane 4 = TGF-β+dh404.
Page 23 of 44
For Peer Review Only
Diabetes
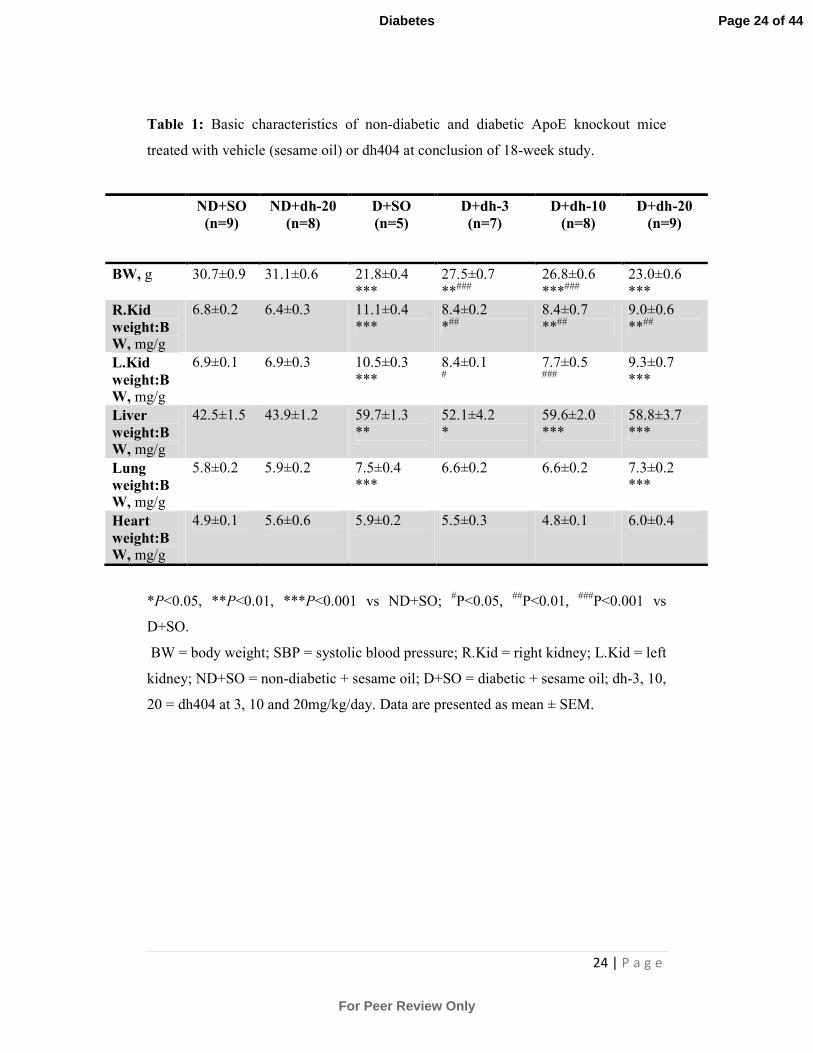
24 | P a g e
Table 1: Basic characteristics of non-diabetic and diabetic ApoE knockout mice
treated with vehicle (sesame oil) or dh404 at conclusion of 18-week study.
ND+SO
(n=9)
ND+dh-20
(n=8)
D+SO
(n=5)
D+dh-3
(n=7)
D+dh-10
(n=8)
D+dh-20
(n=9)
BW, g 30.7±0.9 31.1±0.6 21.8±0.4
***
27.5±0.7
**###
26.8±0.6
***###
23.0±0.6
***
R.Kid
weight:B
W, mg/g
6.8±0.2 6.4±0.3 11.1±0.4
***
8.4±0.2
*##
8.4±0.7
**##
9.0±0.6
**##
L.Kid
weight:B
W, mg/g
6.9±0.1 6.9±0.3 10.5±0.3
***
8.4±0.1 #
7.7±0.5 ###
9.3±0.7
***
Liver
weight:B
W, mg/g
42.5±1.5 43.9±1.2 59.7±1.3
**
52.1±4.2
*
59.6±2.0
***
58.8±3.7
***
Lung
weight:B
W, mg/g
5.8±0.2 5.9±0.2 7.5±0.4
***
6.6±0.2 6.6±0.2 7.3±0.2
***
Heart
weight:B
W, mg/g
4.9±0.1 5.6±0.6 5.9±0.2 5.5±0.3 4.8±0.1 6.0±0.4
*P<0.05, **P<0.01, ***P<0.001 vs ND+SO; #P<0.05,
##P<0.01,
###P<0.001 vs
D+SO.
BW = body weight; SBP = systolic blood pressure; R.Kid = right kidney; L.Kid = left
kidney; ND+SO = non-diabetic + sesame oil; D+SO = diabetic + sesame oil; dh-3, 10,
20 = dh404 at 3, 10 and 20mg/kg/day. Data are presented as mean ± SEM.
Page 24 of 44
For Peer Review Only
Diabetes
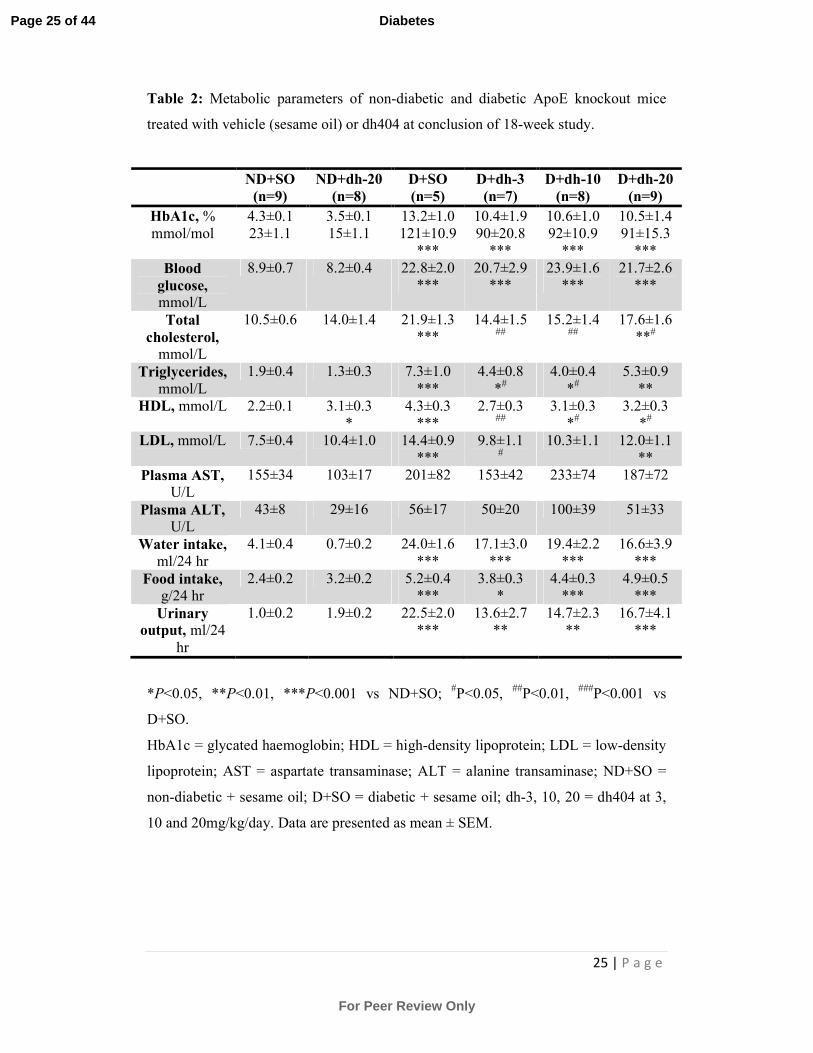
25 | P a g e
Table 2: Metabolic parameters of non-diabetic and diabetic ApoE knockout mice
treated with vehicle (sesame oil) or dh404 at conclusion of 18-week study.
ND+SO
(n=9)
ND+dh-20
(n=8)
D+SO
(n=5)
D+dh-3
(n=7)
D+dh-10
(n=8)
D+dh-20
(n=9)
HbA1c, %
mmol/mol
4.3±0.1
23±1.1
3.5±0.1
15±1.1
13.2±1.0
121±10.9
***
10.4±1.9
90±20.8
***
10.6±1.0
92±10.9
***
10.5±1.4
91±15.3
***
Blood
glucose,
mmol/L
8.9±0.7 8.2±0.4 22.8±2.0
***
20.7±2.9
***
23.9±1.6
***
21.7±2.6
***
Total
cholesterol,
mmol/L
10.5±0.6 14.0±1.4 21.9±1.3
***
14.4±1.5 ##
15.2±1.4 ##
17.6±1.6
**#
Triglycerides,
mmol/L
1.9±0.4 1.3±0.3 7.3±1.0
***
4.4±0.8
*#
4.0±0.4
*#
5.3±0.9
**
HDL, mmol/L 2.2±0.1 3.1±0.3
*
4.3±0.3
***
2.7±0.3 ##
3.1±0.3
*#
3.2±0.3
*#
LDL, mmol/L 7.5±0.4 10.4±1.0 14.4±0.9
***
9.8±1.1 #
10.3±1.1 12.0±1.1
**
Plasma AST,
U/L
155±34 103±17 201±82 153±42 233±74 187±72
Plasma ALT,
U/L
43±8 29±16 56±17 50±20 100±39 51±33
Water intake,
ml/24 hr
4.1±0.4 0.7±0.2 24.0±1.6
***
17.1±3.0
***
19.4±2.2
***
16.6±3.9
***
Food intake,
g/24 hr
2.4±0.2 3.2±0.2 5.2±0.4
***
3.8±0.3
*
4.4±0.3
***
4.9±0.5
***
Urinary
output, ml/24
hr
1.0±0.2 1.9±0.2 22.5±2.0
***
13.6±2.7
**
14.7±2.3
**
16.7±4.1
***
*P<0.05, **P<0.01, ***P<0.001 vs ND+SO; #P<0.05,
##P<0.01,
###P<0.001 vs
D+SO.
HbA1c = glycated haemoglobin; HDL = high-density lipoprotein; LDL = low-density
lipoprotein; AST = aspartate transaminase; ALT = alanine transaminase; ND+SO =
non-diabetic + sesame oil; D+SO = diabetic + sesame oil; dh-3, 10, 20 = dh404 at 3,
10 and 20mg/kg/day. Data are presented as mean ± SEM.
Page 25 of 44
For Peer Review Only
Diabetes
180x204mm (300 x 300 DPI)
Page 26 of 44
For Peer Review Only
Diabetes
180x142mm (300 x 300 DPI)
Page 27 of 44
For Peer Review Only
Diabetes
180x168mm (300 x 300 DPI)
Page 28 of 44
For Peer Review Only
Diabetes

180x139mm (300 x 300 DPI)
Page 29 of 44
For Peer Review Only
Diabetes
180x220mm (300 x 300 DPI)
Page 30 of 44
For Peer Review Only
Diabetes
180x226mm (300 x 300 DPI)
Page 31 of 44
For Peer Review Only
Diabetes

1 | P a g e
SUPPLEMENTARY SECTION
A derivative of Bardoxolone methyl, dh404, in an inverse dose-dependent manner, lessens
diabetes-associated atherosclerosis and improves diabetic kidney disease.
Sih Min Tan1, Arpeeta Sharma
1, Nada Stefanovic
1, Derek Y.C. Yuen
1, Tom C. Karagiannis
2,
Colin Meyer3, Keith W. Ward
3, Mark E. Cooper
1, Judy B. de Haan
1
1Oxidative Stress Laboratory, Diabetic Complications Division, Baker IDI Heart and
Diabetes Institute, Melbourne, Australia 2 Epigenomic Medicine, Baker IDI Heart and Diabetes Institute, Melbourne, Australia
3Reata Pharmaceuticals Inc. 2801 Gateway Dr, Irving, TX 75063, United States.
Running title: dh404 improves end-organ injury in diabetic mice
Corresponding author:
Dr Judy B. de Haan
Address: 75 Commercial Road, Melbourne, Victoria 3004, Australia
Fax number: +61(3) 8532 1100
Phone number: +61(3) 8532 1520
Email address: [email protected]
Page 32 of 44
For Peer Review Only
Diabetes

2 | P a g e
Figure legends
Supplementary Figure 1:
Structural representation of the parent compound Bardoxolone methyl (BM) and its
derivative, dh404, showing the –CF3 modification, an important substitution where the
fluorine on dh404 provides protection from possible rodent-specific toxic metabolites.
Previously it has been shown that agents, BM and dh404, exhibit virtually identical
qualitative properties (that is, dh404 is a reliable surrogate for bardoxolone methyl activity
[1,2]), although dh404 is ~15-fold less potent. Therefore the effects seen in these studies
with dh404 could be expected to have been observed at ~15-fold lower doses of BM, if it
could have been safely dosed to rodents. This makes the pharmacology observed even more
translationally relevant, as it would put therapeutic concentrations in the same range as that
observed in CKD patients with BM.
Supplementary Figure 2: Photomicrographs of adherent NRK cells after exposure to
increasing concentrations of dh404 for 48 and 120h.
NRK cells were treated with dh404 as shown, and allowed to grow for 48 and 120hr in the
presence of the drug. Thereafter, viable cells were assessed by trypan blue exclusion staining
and counting. Significant cell loss was observed after 48h of 1µM dh404. In subsequent
experiments a concentration range of 0 - 0.75µM dh404 for 72h was selected.
Supplementary Figure 3: Immunostaining and quantitation of 4-HNE in the aortic
vessel wall:
Aortic sections were immunostained (brown staining) with an antibody against 4-HNE to
detect lipid oxidative damage within the vessel wall. Representative photomicrographs are
shown in (A). Quantitation of 4-HNE staining is shown in (B). A trend towards an increase in
4-HNE was observed in diabetic aortas treated with sesame oil and this trend was reversed in
the presence of 3mg/kg dh404, albeit that this failed to reach significance. ND = non-diabetic
mice; SO = sesame oil; dh-3,-10,-20 = dh404 at 3, 10 or 20mg/kg/day; D = diabetic mice.
Supplementary Figure 4: Immunostaining and quantitation of 4-HNE in aortic plaque:
Aortic sections were immunostained (brown staining) with an antibody against 4-HNE to
detect lipid oxidative damage within the plaque. Representative photomicrographs are shown
in (A). Quantitation of 4-HNE staining is shown in (B). There was a diabetes-associated
increase in 4-HNE staining in the diabetic plaque (although this failed to reach significance
Page 33 of 44
For Peer Review Only
Diabetes

3 | P a g e
by one-way ANOVA and post hoc testing). This trend was reversed at higher doses of dh404,
albeit that this failed to reach significance. ND = non-diabetic mice; SO = sesame oil; dh-3,-
10,-20 = dh404 at 3, 10 or 20mg/kg/day; D = diabetic mice.
Supplementary Figure 5: Immunostaining and quantitation of nitrotyrosine (NT) in
aortic plaque:
Aortic sections were immunostained (brown staining) with an antibody against NT to detect
protein oxidative damage within the plaque. Representative photomicrographs are shown in
(A). Quantitation of NT staining is shown in (B). There was a diabetes-associated increase in
NT staining in the diabetic plaque (although this failed to reach significance by one-way
ANOVA and post hoc testing). This trend was reversed at higher doses of dh404, albeit that
this failed to reach significance. ND = non-diabetic mice; SO = sesame oil; dh-3,-10,-20 =
dh404 at 3, 10 or 20mg/kg/day; D = diabetic mice.
Supplementary Figure 6: Analysis of tubulointertitial injury
Kidney sections were subjected to immunohistochemical staining with an antibody against
collagen IV. Representative photomicrographs are shown in (A). Quantitation of collagen IV
staining is shown in (B). There was a trend towards an increase in collagen IV staining in
diabetic kidney tubules and this was reversed after 3mg/kg/day dh404 treatment for 18
weeks. 10- and 20mg/kg/day dh404 did not appear to reduce this trend. ND = non-diabetic
mice; SO = sesame oil; dh-3,-10,-20 = dh404 at 3, 10 or 20mg/kg/day; D = diabetic mice.
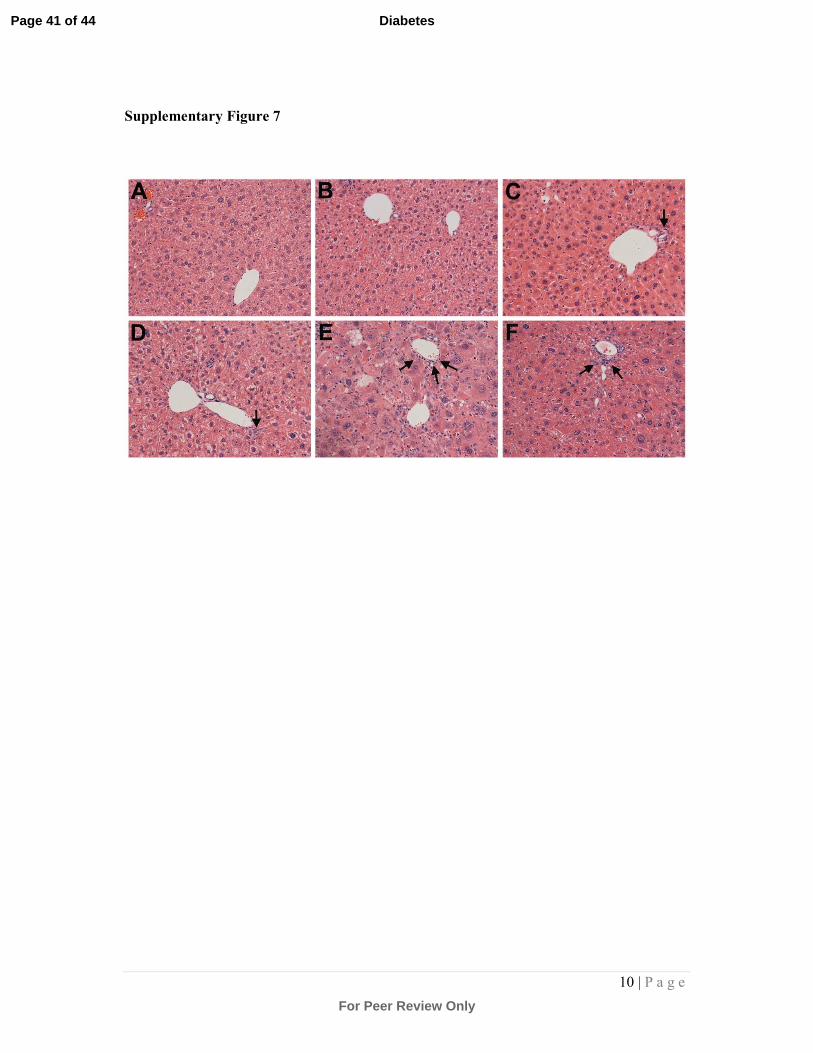
Supplementary Figure 7: Photomicrographs of liver sections stained with haematoxylin
and eosin:
Liver sections were stained with haematoxylin and eosin to visualise hepatocytes and
inflammatory cells enabling both the semi-quantitation of hepatocellular cytoplasmic
rarification, and the extent of inflammatory infiltration from the portal vein. (A) Non-diabetic
ApoE-/- mouse liver; (B) Non-diabetic ApoE-/- mouse liver treated with 20mg/kg dh404;
(C) Diabetic ApoE-/- mouse liver treated with SO; (D) Diabetic ApoE-/- mouse liver treated
with 3mg/kg dh404; (E) Diabetic ApoE-/- mouse liver treated with 10mg/kg dh404; (F)
Diabetic ApoE-/- mouse liver treated with 20mg/kg dh404. Arrows point to infiltrating
inflammatory cells surrounding the portal vein.
Page 34 of 44
For Peer Review Only
Diabetes
4 | P a g e
Supplementary Figure 1
Page 35 of 44
For Peer Review Only
Diabetes

5 | P a g e
Supplementary Figure 2
Page 36 of 44
For Peer Review Only
Diabetes
6 | P a g e
Supplementary Figure 3
Page 37 of 44
For Peer Review Only
Diabetes
7 | P a g e
Supplementary Figure 4
Page 38 of 44
For Peer Review Only
Diabetes
8 | P a g e
Supplementary Figure 5
Page 39 of 44
For Peer Review Only
Diabetes
9 | P a g e
Supplementary Figure 6
Page 40 of 44
For Peer Review Only
Diabetes
10 | P a g e
Supplementary Figure 7
Page 41 of 44
For Peer Review Only
Diabetes
11 | P a g e
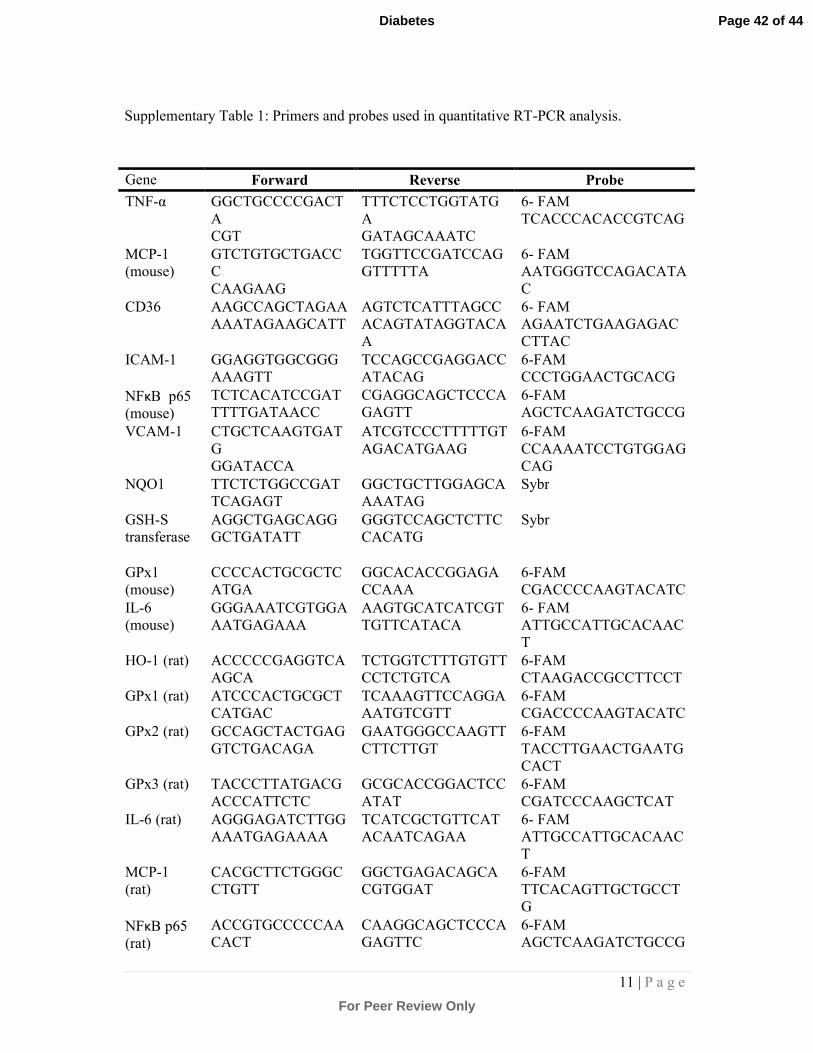
Supplementary Table 1: Primers and probes used in quantitative RT-PCR analysis.
Gene Forward Reverse Probe
TNF-α GGCTGCCCCGACT
A
CGT
TTTCTCCTGGTATG
A
GATAGCAAATC
6- FAM
TCACCCACACCGTCAG
MCP-1
(mouse)
GTCTGTGCTGACC
C
CAAGAAG
TGGTTCCGATCCAG
GTTTTTA
6- FAM
AATGGGTCCAGACATA
C
CD36 AAGCCAGCTAGAA
AAATAGAAGCATT
AGTCTCATTTAGCC
ACAGTATAGGTACA
A
6- FAM
AGAATCTGAAGAGAC
CTTAC
ICAM-1 GGAGGTGGCGGG
AAAGTT
TCCAGCCGAGGACC
ATACAG
6-FAM
CCCTGGAACTGCACG
NFκB p65
(mouse)
TCTCACATCCGAT
TTTTGATAACC
CGAGGCAGCTCCCA
GAGTT
6-FAM
AGCTCAAGATCTGCCG
VCAM-1 CTGCTCAAGTGAT
G
GGATACCA
ATCGTCCCTTTTTGT
AGACATGAAG
6-FAM
CCAAAATCCTGTGGAG
CAG
NQO1 TTCTCTGGCCGAT
TCAGAGT
GGCTGCTTGGAGCA
AAATAG
Sybr
GSH-S
transferase
AGGCTGAGCAGG
GCTGATATT
GGGTCCAGCTCTTC
CACATG
Sybr
GPx1
(mouse)
CCCCACTGCGCTC
ATGA
GGCACACCGGAGA
CCAAA
6-FAM
CGACCCCAAGTACATC
IL-6
(mouse)
GGGAAATCGTGGA
AATGAGAAA
AAGTGCATCATCGT
TGTTCATACA
6- FAM
ATTGCCATTGCACAAC
T
HO-1 (rat) ACCCCCGAGGTCA
AGCA
TCTGGTCTTTGTGTT
CCTCTGTCA
6-FAM
CTAAGACCGCCTTCCT
GPx1 (rat) ATCCCACTGCGCT
CATGAC
TCAAAGTTCCAGGA
AATGTCGTT
6-FAM
CGACCCCAAGTACATC
GPx2 (rat) GCCAGCTACTGAG
GTCTGACAGA
GAATGGGCCAAGTT
CTTCTTGT
6-FAM
TACCTTGAACTGAATG
CACT
GPx3 (rat) TACCCTTATGACG
ACCCATTCTC
GCGCACCGGACTCC
ATAT
6-FAM
CGATCCCAAGCTCAT
IL-6 (rat) AGGGAGATCTTGG
AAATGAGAAAA
TCATCGCTGTTCAT
ACAATCAGAA
6- FAM
ATTGCCATTGCACAAC
T
MCP-1
(rat)
CACGCTTCTGGGC
CTGTT
GGCTGAGACAGCA
CGTGGAT
6-FAM
TTCACAGTTGCTGCCT
G
NFκB p65
(rat)
ACCGTGCCCCCAA
CACT
CAAGGCAGCTCCCA
GAGTTC
6-FAM
AGCTCAAGATCTGCCG
Page 42 of 44
For Peer Review Only
Diabetes

12 | P a g e
col I (rat) TGCCGATGTCGCT
ATCCA
TCTTGCAGTGATAG
GTGATGTTCTG
6- FAM
CCTTCCTGCGCCTGA
col IV (rat) CACTATGAAAACC
GTAAAGTGCCTTA
GCAAACAGAGGCC
AACGAA
6- FAM
ATTTGCGTAACTAACA
CACC
Fn (rat) CATGGCTTTAGGC
GAACCA
CATCTACATTCGGC
AGGTATGG
6- FAM
CCCCGTCAGGCTTA
18S TGTTCACCATGAG
GCTGAGATC
TGGTTGCCTGGGAA
AATCC
6-FAM
TGCTGGCACCAGACCA
GACTTGCCCTC
Page 43 of 44
For Peer Review Only
Diabetes
13 | P a g e
Supplementary Table 2: Histopathological assessment of hepatocellular cytoplasmic
rarification in liver sections.
Treatment
Group
Number of
livers examineda
Hepatocellular
cytoplasmic rarification
score (0-5) b
Chronic periportal
inflammation score (0-5) c
ND 8 0.61 ± 0.15 0.21 ± 0.06
ND+dh-20 8 1.10 ± 0.27 0.29 ± 0.04
D+SO 5 0.59 ± 0.13 0.46 ± 0.08
D+dh-3 7 1.26 ± 0.36 0.48 ± 0.13
D+dh-10 7 1.33 ± 0.27 0.87 ± 0.23*
D+dh-20 8 0.70 ± 0.29 0.68 ± 0.2
*P<0.05 vs ND.
ND = non-diabetic; D+SO = diabetic + sesame oil; dh-3, 10, 20 = dh404 at 3, 10 and
20mg/kg/day. Data are presented as mean ± SEM. a
For each liver, 10 sections were quantitated and averaged. b To determine hepatocellular cytoplasmic rarification, the following scale was used: 0 =
normal pathology with minor lipid inclusions (normal pathology for ApoE-/- livers);
1=minimal change; 2=mild change; 3=moderate change; 4=marked change; 5=severe change
as per Chin et al.[2] c
To determine periportal inflammation, the following scale was used: 0 = minimal
infiltration (<10 infiltrating inflammatory cells in the area surrounding the portal vein);
1=minimal change (<50 infiltrating inflammatory cells); 2=mild change (50-75 inflammatory
cells); 3=moderate change (75-100 inflammatory cells); 4=marked change (100-150
inflammatory cells); 5=severe change (>150 inflammatory cells).
REFERENCES
1. Ichikawa T, Li J, Meyer CJ, Janicki JS, Hannink M, et al. (2009) Dihydro-CDDO-
trifluoroethyl amide (dh404), a novel Nrf2 activator, suppresses oxidative stress in
cardiomyocytes. PLoS One 4: e8391.
2. Chin M, Lee CY, Chuang JC, Bumeister R, Wigley WC, et al. (2013) Bardoxolone Methyl
Analog Rta 405 and Dh404 Are Well-Tolerated and Exhibit Efficacy in Rodent
Models of Type 2 Diabetes and Obesity. Am J Physiol Renal Physiol 304: F1438-
1446.
Page 44 of 44
For Peer Review Only
Diabetes


















